HISTOLOGIE
nauka o tkáních
Materiály slouží ke studiu daného předmětu.
Jiné
použití konzultujte laskavě s autorem.
(počet přístupů od 15.3.07)
Kontakt
na učitele: ptacek@sci.muni.cz
Anglicko český slovník pro
případnou potřebu překladu při studiu převzatých obrázků:
http://www.translator.cz/bin/translator
|
Kapitoly které v obsahu nejsou aktivovány jsou pouze o něco kratší, nikoli méně významné! 2.
Vznik tkání 2.1.
Rýhování
zygoty při pohlavním rozmnožování 2.2.
Gastrulace a histogeneze některých bezobratlých 2.3.
Typy tělní dutiny 2.4.
Rozdělení organismů podle
složitosti tělesné organizace 2.5.
Diferenciace
tkání 2.6.
Tkáňové buňky 2.7.
Původ tkání 3.
DRUHY
TKÁNÍ 4.
Epitely 4.1.
Dělení epitelů
podle počtu vrstev 4.2.
Dělení epitelů
podle tvaru buněk 4.3.
Dělení epitelů podle polárního rozlišení 4.4.
Dělení epitelů podle funkce 4.4.1.
Krycí epitely 4.4.2.
Výstelkové
epitely 4.4.3.
Žlázové
epitely 4.4.4.
Resorbční epitely 4.4.5.
Smyslové buňky 4.4.6.
Zárodečné
epitely 4.4.7.
Svalové epitely 4.4.8.
Pigmentové
epitely 4.5.
Ontogeneze epitelů 5.
POJIVA 5.1.
Struktura
pojiv 5.1.1.
Pevné buňky (fixní) 5.1.2.
Volné
buňky 5.1.3.
Mezibuněčné hmoty 5.2.
Druhy pojiv 5.3.
Mezoglea 5.4.
Zárodečný mezenchym 5.5.
Rosolovité pojivo 5.6.
Retikulární (síťovité) pojivo 5.7.1.
Fixní buňky řídkého vláknitého pojiva 5.7.2.
Volné buňky řídkého vláknitého pojiva 5.8.
Tukové pojivo
5.9.
Tuhé vláknité pojivo – vazivo 5.9.1. Plsťovité
vazivo 5.9.2.
Provazcovité vazivo 5.9.3.
Vaziva elastická
5.10. Pojivo buněčné
5.11. Pojiva chrupavková 5.12. Kost 5.12.1. Kost primární (vláknitá) 5.12.2. Kost definitivní (lamelózní) 5.12.3. Osifikace 5.12.4. Spoje kostrové 5.13. Zubní tkáně
5.14. Pojiva trofická (tělní tekutiny) 5.14.1. Míza 5.14.2. Krev 5.14.3. Teorie vzniku krevních buněk 5.14.4. Tkáňový mok 5.14.5.
Mozkomíšní
mok 5.14.6.
Krvomíza
– hemolymfa u hmyzu 5.14.7.
Hemocyty hmyzu 5.14.8.
Krevní barviva 6.1. Myoepitely 6.2. Hladká
svalovina 6.3. Žíhaná svalovina 6.3.1. Elektrická tkáň 6.4.
Myokard 7.1. Základní
systémy nervové tkáně 7.4.
Diferenciace nervové tkáně 8.1.
Typy gamet 8.2.
Vznik gamet 10. Použitá literatura |
Zde najdete stručný sylabus bez obrázků,
vhodný pro tisk
1. Charakteristika živočicha
Těla mnohobuněčných živočichů tvoří různé soustavy
buněk diferencovaných (doslova rozrůzněných) k určitému úkolu. Specializace
pokročila natolik, že buňky ztrácejí schopnost samostatného života. Jen u
některých primitivních organismů (Porifera) (URL
1) je takový návrat ještě možný. Specializace i diferenciace roste spolu
s růstem úrovně tzv. fylogenetického vývoje.
Jen některé buňky (určené k rozmnožování
organismu nebo jeho buněk) mohou existovat určitou dobu samostatně a podržet si schopnost dát vznik buď celému
organismu (tzv. totipotentní - všeho schopné blastomery raných stadií
embryonálního vývoje) nebo určitým
typům buněk tkáňových (tzv. pluripotentní - mnohého schopné zvané též
jako kmenové buňky). Organismy si většinou udržují jakousi banku buněk prapohlavních,
z nichž redukčním dělením vznikají buňky pohlavní. O všechny uvedené typy
buněk většinou pečují buňky jiné.
Činnost buněk v soustavách je koordinována a
regulována opět soustavami buněk jiných (nervové a žlázové buňky). V důsledku
vnitřní koordinace a regulace životních pochodů tvoří mnohobuněčné organismy
jediný harmonický celek - živočišného jedince.
|
Připomeňme
si zde znova charakteristiku živočicha, z níž se jednotlivosti
v diferenciaci buněk do tkání odvíjejí: Živočich je eukaryotický
mnohobuněčný heterotrofní organismus, v jehož vývoji se vyskytují
nejméně dvě vrstvy buněk (stadium gastruly) -Buňky jsou
diferencovány k určitým úkolům do souborů (tkání) -Tkáně mohou
vytvářet orgány nebo soustavy orgánů zabezpečujících životní pochody a funkce
živočicha, které spolu logicky souvisejí: -Příjem a
zpracování organické potravy (nutnost potravu v prostředí objevit,
proto se vytvořily:) -Dráždivost a
orientace v prostředí (registrace potravy, partnera, nepřítele …
vyžaduje:) -Schopnost
pohybu buď celého živočicha nebo jeho částí (Vlastnost pro živočichy
charakteristická. Nepohybuje –li se živočich, pohybuje se prostředí, ve
kterém žije) -Vyšší
nervová a duševní činnost (Není dána všem živočichům a zřejmě se
nevyskytuje u jiných forem organismů.) -Složité
vnitřní členění jak co do počtu buněk a jejich typů, tak i co do rozmanitosti
tkání a orgánů. Povrch často jednoduchý. -Schopnost
reprodukce (Není výlučnou vlastností živočichů – je charakteristická pro
všechny organismy) |
2. vznik tkání
Tkáně (URL 20) se
diferencují a specializují postupně (v čase) během ontogenetického vývoje. Původně
většinou z jediné buňky při pohlavním rozmnožování (nebo skupiny buněk při
rozmnožování nepohlavním) se vytvářejí dělením a uspořádáním (polohou) první
tzv. zárodečné listy, ze kterých se postupně diferencuje další tkáň.
![]()
2.1. Rýhování zygoty při pohlavním
rozmnožování
Při pohlavním
rozmnožování vzniká splynutím vajíčka a spermie tzv. zygota.
Splynutím jader vzniká pronukleus (samčí proteiny
sbalující DNA jsou nahrazeny histony vajíčka).
Následuje prudký pokles entropie zygoty a posléze
její dělení – rýhování vajíčka.
Vznikající dceřinné buňky –
blastomery (jsou většinou totipotentní – schopné dát vznik celému organismu)
Zygota - 2
blastomery - 4 blastomery
(totipotence buněk většinou zachována),
8 blastomer,…a dále … totipotence
se ztrácí.
Morula –
nahloučení buněk vznikajícího zárodku do skupiny bez vnitřní dutiny (plod
moruše)
Blastula –
tvar s dutinou uprostřed (blastocoel – prvotní dutina tělní), blastoderm
(první tkáň epitelového typu).
Gastrula –
vznik dvojvrstevného organismu (invaginace, imigrace, delaminace, epibolie). Na
gastrule rozeznáváme:
ektoderm (-blast)
entoderm (-blast)
blastocoel (prvotní dutina tělní)
blastoporus (prvoústa)
archenteron (prvostřevo)
(Prvoústí – blastoporus zůstává ústy a
vylučovací otvor se tvoří druhotně a druhoústí – blastoporus se mění ve
vylučovací otvor a ústa se tvoří druhotně.)
Podle starších teorií byli živočichové děleni na
Diblastica
- živočichové, jejichž vývoj zůstává na úrovni dvou zárodečných listů – ektodermu
a entodermu.
Triblastica
- živočichové, u nichž se postupně vytváří i třetí zárodečný list – mezoderm
(blast.)
Dnes jsou tyto názory opuštěny, protože u řady
živočichů se třetí zárodečný list nezakládá na způsob epitelu, ale jako tkáň,
která vyplňuje prvotní dutinu tělní. (Ta se ovšem v podobě mezoglei
vyskytuje i u houbovců a žahavců.)
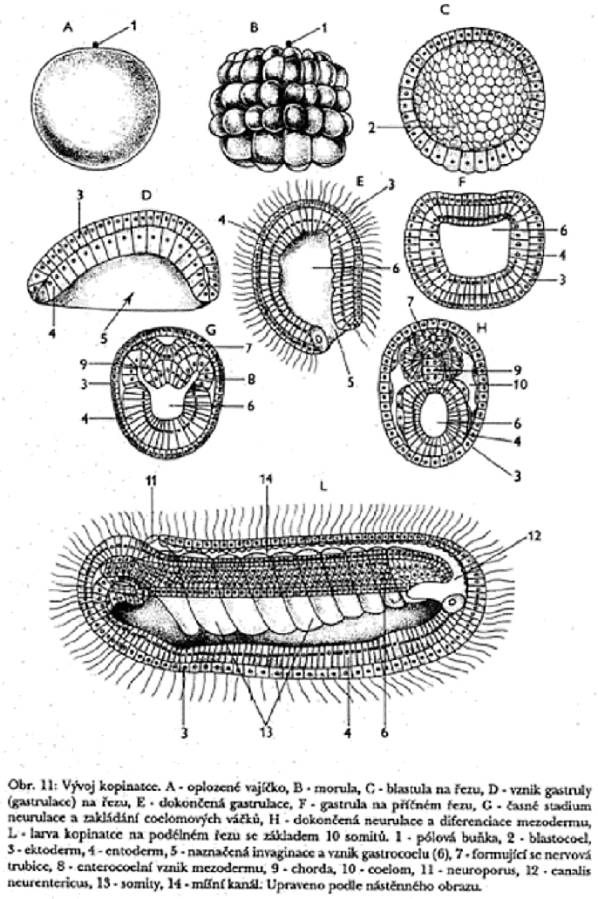
Nahoře - schéma vývoje
kopinatce, enterocoelní (vychlípením ze střeva) vznik coelomových váčků.
Jejich další diferenciací vznikají somit nahoře a splanchnotom dole.
Pro srovnání gastrulace a histogeneze u kopinatce ještě v barvách:
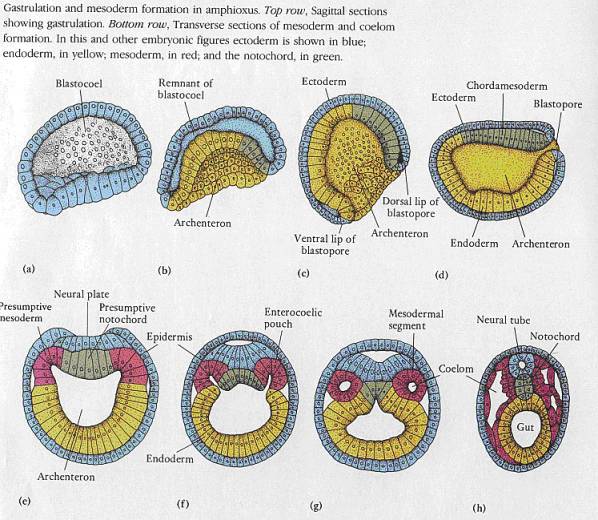
U obratlovců vzniká mezoderm podobně (viz dolní obrázek). Diferenciací
buněk somitu vznikají: dermatom
(škára), myotom (kosterní svalovina), sklerotom (kostra).
Splanchnotom obou stran těla se na ventrální (břišní) straně spojuje v jediný váček a vytváří ventrální úseky druhotné dutiny tělní (u vyšších obratlovců dutinu břišní, hrudní a osrdečníkovou). V těchto dutinách se tzv. somatopleura mění v epitel vystýlající vnější část dutiny (pobřišnice, pohrudnice) a splanchnopleura tvoří mezotel, kryjící vnitřní orgány (játra, žaludek, střeva atp.) nebo tzv. poplicnici, kryjící plíce. Hřbetní část váčků splanchnopleury zůstává zachována a mění se v tzv. mesenterium (střevní závěs, umožňující vaskularizaci (krevní zásobení) a inervaci trávících orgánů.
Pohlavní orgány a ledviny vznikají z dorzální části splanchnotomu.
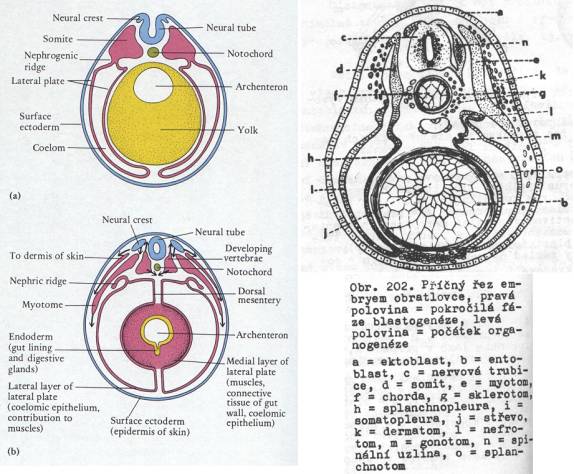
Schéma histogeneze u obratlovců příčný řez
tělem). Modře ektoderm – epidermis a
nervové trubice vzniklá vchlípením ektoblastu na hřbetní straně zárodku.
Z neurálních lišt (neural crest) vznikají např. pigmentové buňky, spinální
ganglia, část zubu, některé kosti viscerálního skeletu a údajně též buňky
Langerhansových ostrůvků v pankreatu. Červeně – mezoderm. Dermatom se
rozpadá a vcestuje pod epidermis jako buňky tvořící škáru. Sklerotom se
vyvine v obratle a kostru, myotom v kosterní svalovinu. Dorzální
mezenterium zůstává, ventrální zaniká a váčky dole splývají. Notochord – struna
hřbetní (u mnohých z entodermu). Yolk – žloutek, budoucí entoderm.
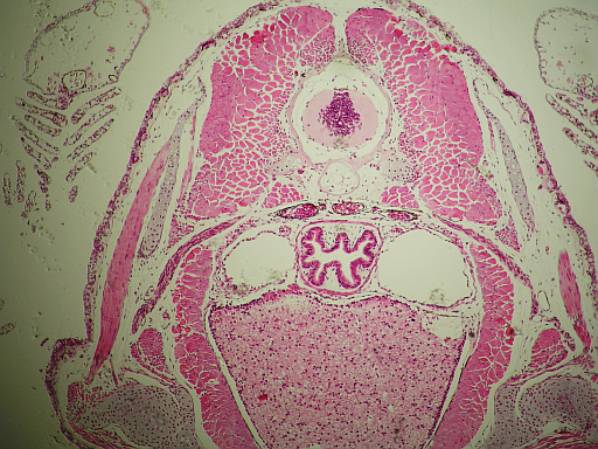
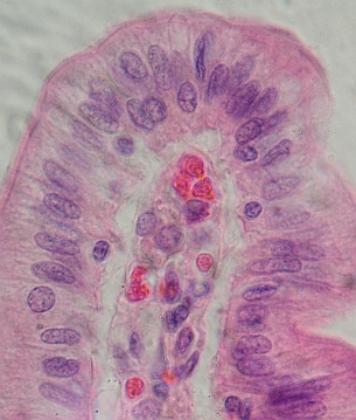
Příčný řez larvou (srovnej též obrázek nahoře) čolka
(Triturus sp.) v oblasti pletence ramenního. Srovnej: epidermis, pod ní
vcestované fibroblasty škáry (dermatom), v dorzální části mohutný myotom
rozpadávající se ve hřbetní svalovinu (červeně). Nahoře uprostřed mícha
s šedou hmotou (těla neuronů) uprostřed, krytá vazivovým pouzdrem, které
se osifikací změní v obratle, po stranách jsou vidět chrupavčité základy
jejich výběžků. Pod míchou je jemně vyvinutá struna hřbetní, která postupně
zanikne. Pod chordou se nachází aorta dorsalis po stranách aorty základy ledvin
(nefrotom). V hrudní dutině (pod chordou) se nachází jícen a po stranách
dvě zatím jednoduché vakovité plíce. Řezem zachycená přední část břišní dutiny
je vyplněna mohutnými játry. Po stranách dutiny břišní jsou základy kosterní
svaloviny pletence předních končetin, jehož chrupavčité základy jsou vidět
v dolních rozích. V horních rozích jsou řezy žábry. Tmavé body až
linie v pojivových obalech vnitřních orgánů a pod epidermis jsou velmi
hojné pigmentové buňky.
![]()
2.2. Gastrulace a histogeneze některých bezobratlých
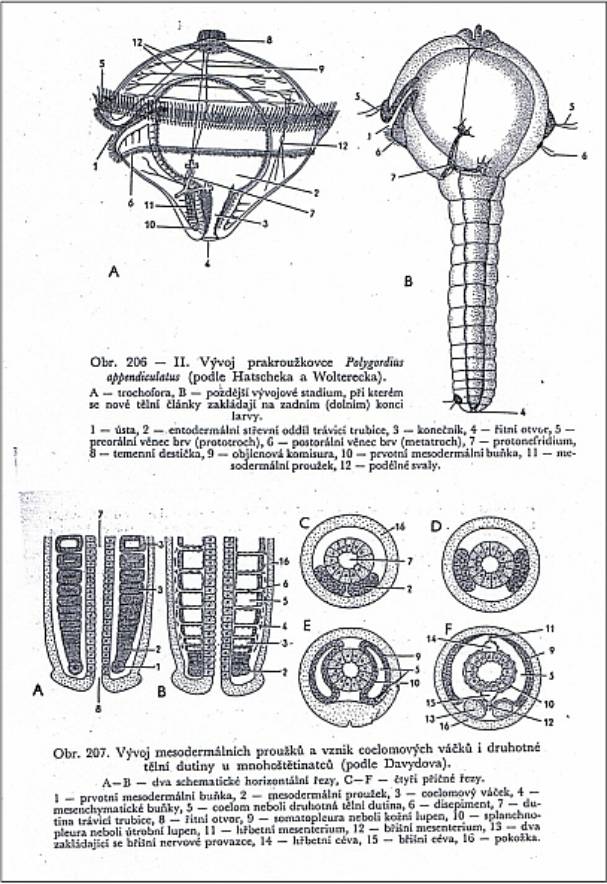
Nahoře
příklad vzniku mezodermu teloblastickým způsobem (kroužkovci). Převzato
z Dogiela (1961). Dva teloblasty (pluripotentní buňky) vycestují do
blastocoelu larvy trochofory a začnou se dělit. Každá z dceřinných buněk
vzniklých ze 2 teloblastů, dává vznik samostatné dutině - coelomovému
váčku. Kroužkovec se tvoří směrem vzad. Jeden kroužek např. žížaly obsahuje
tedy dva váčky. Břišní nervová páska se vchlipuje na ventrální (břišní) straně
těla z ektodermu. Z listů mezodermu vzniká podkožní svalovina,
metanefridie, svalovina, svalovina kolem trávící trubice, chloragogenní tkáň
(vylučování), cévní soustava. Na posledním obrázku vpravo dole je dobře
zřetelné, že dutina cévní soustavy (14) je zbytkem prvotní dutiny tělní.
![]()
2.3. Typy tělní
dutiny

Struktura
kroužkovce (podle Barnes a kol.)
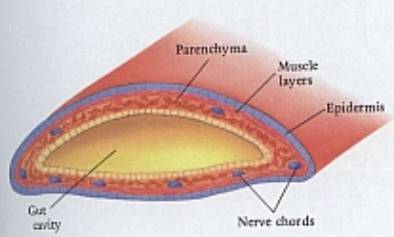
Schéma
tělní dutiny typu schizocoelu (ploštěnci), jenž je rozptýlen v parenchymu
střední vrstvy tělních buněk.
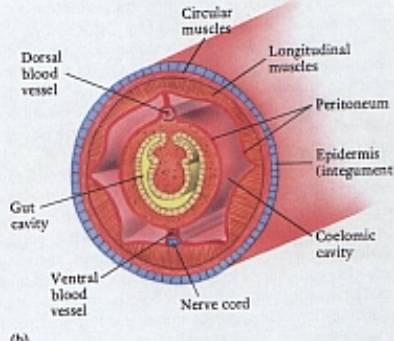
Schéma
druhotné tělní dutiny - coelomu
kroužkovce.
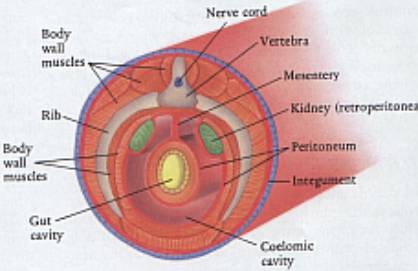
Schéma
tělní dutiny (coelomu) obratlovce.
Dalším
typem tělní dutiny je pseudocoelom (viz obr. dole) – mají ji organismy
s podobnou tělní stavbou jako hlístice (Nematoda). Představuje volný
prostor kolem trávící trubice.
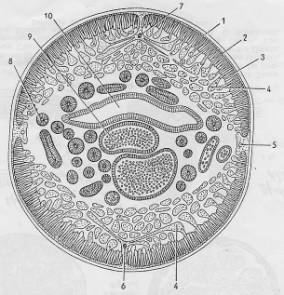

Konečně,
tělní dutina členovců se nazývá mixocoel. Představuje směs prvotní a
druhotné dutiny a vzniká rozpadem zárodečných mezodermu během embryogeneze.
Nachází se u členovců.
Souborně všechny tkáně vývoje z: označujeme
jako:
ektoblastu ektoderm
entoblastu entoderm
mezoblastu mezoderm
(mezenchym) - vycestovaný materiál do dutin mezi
zárodečnými listy.
Vývoj mezodermu, jak již bylo uvedeno na obrázcích,
může probíhat např. enterocoelně nebo teloblasticky:
teloblasticky – ze dvou teloblastů
vycestovaných do blastocoelu - např. Annelida -kroužkovci
enterocoelně
- z entodermu Branchiostoma (kopinatec) .
![]()
2.4. Rozdělení organismů
podle složitosti tělesné organizace
Diblastica - dvojvrstevní - mají pouze dvě
vrstvy buněk - ektoderm a entoderm:
Placozoa - vločkovci
Porifera - houby
Cnidaria - žahavci
Ctenophora - žebernatky
Dyciemida
Orthonectida
Triblastica - trojvrstevní - mají třetí vrstvu buněk
Ta může vzniknout z hvězdicovitých buněk
vycestovaných během embryonálního vývoje do blastocoelu, tzv. mesenchymu.
Sem lze zařadit např. následující kmeny:
Platyhelmintes - ploštěnci
Rotifera – vířníci
Chaetognatha – ploutvenky
Gastrotricha - břichobrvky
Nematoda - hlístice
Nematomorpha - strunovci
Cephalorhyncha - chobotovci
Acanthocephala - vrtejši
Triblastica, praví trojvrstevní - v době
zárodečného vývoje vytvářejí pravý zárodečný list a coelomové váčky, které se postupně mohou opět
rozpadnout.
Nemertini - pásnice
Mollusca - měkkýši
Sipunculidea - sumýšovci
Echiuridea - rypohlavci
Annelida - kroužkovci
Pogonophora - vláknonoši
Tardigrada - želvušky
Arthropoda - členovci
Tentaculata - chapadlovci
Entoprocta - mechovnatci
Priapulidea - hlavatci
Loricifera
Cycliophora
Hemichordata - polostrunatci
Chordata - strunatci
![]()
2.5 Diferenciace tkání
- specializace buněk k určité činnosti –
nastává postupně v procesu histogeneze (vzniku tkání) a organogeneze
(vzniku orgánů)
I. Vysvětlení pomocí (embryonální) indukce:
Buňky vylučují během své existence
látky, které mají schopnost ovlivnit aktivitu genů buněk sousedních. Tím buňky,
které již diferenciaci prodělaly, mohou indukovat diferenciaci jiných buněk
(částečně prokázáno, ale ne vznik tvarů, např. rozdílný tvar ruka – noha při
stejných genech).
II. Působení morfogenetického pole:
Způsob
nabývání tvaru (uspořádání buněk v prostoru i s časovým průběhem) je
z dřívějších podobných pochodů zaznamenám v informačních polích mimo
organismus, který z tohoto záznamu čerpá (dosud neprokázáno vědecky – teorie morfické rezonance, Dr. R. Sheldrake).
2.6. Tkáňové buňky
Stavba tkání je rozmanitá podle účelu. Často se na ní
účastní několik typů buněk:
- buňky fixní - pevné – nepohyblivé, určující
pro typ tkáně.
- buňky volné - pohyblivé: aktivně
(monocyt),pasivně (erytrocyt)
Často více nebo méně mezibuněčné hmoty
(tekuté, polotekuté, pevné nebo dosedají buňky těsně na sebe).
Buňky fixní mohou být spojeny výběžky, které
prostupují mezibuněčnou hmotou a propojují buňky dosti vzdálené. Např.
embryonální mesenchym, kostní tkáň. (Takové spoje jsou běžné i u rostlin, kde
se nazývají - plazmodezmy
URL 5.)
Spoje mezi buňkami ve tkáních s malým množstvím
mezibuněčných hmot: interdigitace
(URL 2), dezmozómy
(s využitím bílkovinných vláken cytoskeletu - tonofibrilami), terminální
uzávěry (čočkovité ztluštěniny bez vláken), nebo přímý kontakt membrán.
Plazmodezma
(URL 6) je vlastně kanál mezi dvěma přilehlými buňkami, umožňující výměnu
cytoplazmy mezi nimi. Plazmatická membrána jedné buňky je kontinuálně propojena
s plazmatickou membránou druhé buňky. Většinou mají ve svém středu úzké
válcovité dezmotubuly, odvozené z endoplazmatického retikula, které se zdají
být propojeny s endoplazmatickými retikuly obou buněk. Mezi dezmotubuly a
plazmatickou membránou se nachází úzký prstenec cytoplazmy zvaný annulus.
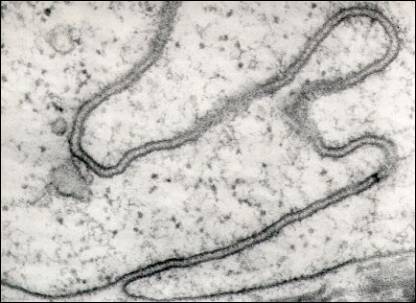
Interdigitace
(URL 3)
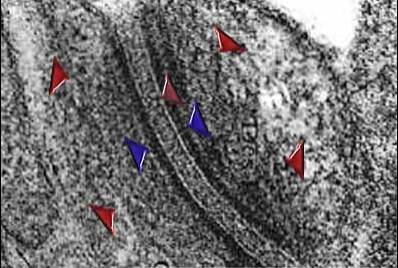
Dezmozómy
(URL 4) jsou komplexy, které spojují protilehlé plazmatické membrány. Každý
dezmozóm se skládá z destičky o vysoké elektronové hustotě nacházející se
těsně pod plazmatickou membránou (modré šipky), ke které se sbíhají
tonofilamenta (červené šipky). Prostor mezi plazmatickými membránami, široký
obvykle asi 25 nm, je vyplněn elektronově hustým materiálem (hnědá šipka)
Pouta: chemické vazby, elektrické síly, koheze
(fyzikálně-chemické síly)
Syncytia -
rozrušením sousedních membrán buňky splývají - usnadňuje se mezibuněčný
transport, odvod vyrobených látek i celá funkčnost dané tkáně (myokard, žíhaná
svalovina).
Mezibuněčná hmota:
fibrilární - tekuté polotekuté, pevné,
interfibrilární (amorfní) různé organické i
anorganické látky.
![]()
2.7. Původ tkání
Buňky podle zárodečných listů: (URL 7) 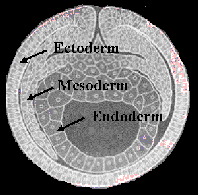
Ektoderm:
Buňky tělního pokryvu a výstelkové epitely, včetně z nich
diferencovaných žláz, případně respiračních epitelů ektodermálních dýchacích
orgánů, neurony (nervové buňky), gliové buňky, pigmentové buňky, údajně i buňky
Langerhansových ostrůvků slinivky břišní), buňky Malphigických žláz hmyzu, tracheální systém hmyzu, pigmentové buňky,
rohovka a čočka oka
Mezoderm:
Mezenchym – nepravidelně
uspořádané buňky vycestované do prvotní tělní dutiny z ektodermu (ektomezenchym
– vzácnější) nebo entodermu (entomezenchym – častější). Z nich
se tvoří např. vnitřní orgány primitivních (tzv. primárních) larev.
Z pravého třetího zárodečného listu – mezoblastu
(mezodermu) vznikají:
přes tzv. zárodečný mezenchym –
reticuloblast – retikulární buňka (nikoli logicky
retikulocyt - termín obsadili hematologové))
fibroblast – fibrocyt (produkce mezibuněčných hmot ve
vláknitých pojivech)
lipoblast – adipocyt (tukové pojivo)
chondroblast – chondrocyt (chrupavky)
osteoblast – osteocyt (kostní tkáně)
pluripotentní buňky krvetvorby
myoblast – sarkocyt (buňka hladké svaloviny)
myoblast(-ty) – myotuby – svalové vlákno (žíhané
svaloviny)
endotel cév
peritoneum (mezotel)
Nediferencovaný mezenchym – Marchandovy buňky - hojení ran.
K mezodermu patří:
výstelka druhotné dutiny tělní – peritoneum, mezotel – základní buňky pojiv
kromě chordy, svalové buňky, funkční buňky vylučovacích systémů, zárodečné
epitely, krvinky, endotel cév i mízní kapiláry.
Entoderm:
Rezorbční a žlázové epitely střední části trávící soustavy (buňky hlavní, krycí, hlenotvorné v žaludku, enterocyty střeva, Lieberkühnových krypt, exosekretorické buňky pankreatu, hepatocyty, rezorbční epitely plic a žaber obratlovců), chordoblasty a chordocyty struny hřbetní (podmíněně, ne u všech strunatců) buňky Malphigických žláz pavoukovců, chorda dorsalis.
![]()
3.
DRUHY TKÁNÍ (URL 8) podle Köligera:
1. Epitely
(URL 13) (tkáně
epitelové)
2. Pojiva (tkáně pojivové)
3. Tkáně svalové (svalovina)
4. Tkáně nervové
5. Pohlavní buňky
4. Epitely
Fylogeneticky i ontogeneticky nejpůvodnější tkáně
rozprostřené do plochy z jedné nebo více vrstev buněk - něco kryjí nebo
vystýlají (buňky fixní). Jsou to jen fixní buňky, téměř bez mezibuněčné
hmoty (pokud je, pak je tvořena převážně kyselinou hyaluronovou). Buňky
nasedají těsně na sebe. Pod vrstvou buněk a sousední tkání vrstva kolagenních a
retikulárních vláken v tenké dvojvrstvě = bazální lamina. Vnější strana vždy
obnažena - tvoří povrch těla nebo vnější či vnitřní povrch orgánů.
Polární rozlišení epitelů: svrchní, apikální vrstva se liší od bazální. Apikální pól -
funkční, bazální pól - upevnění, regenerace, předávání látek.

(URL
10)
Dělení
epitelů:
a) podle počtu vrstev,
b) podle tvaru buněk,
c) podle polárního rozlišení,
d) podle funkce
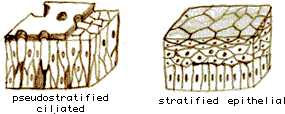
4.1. Dělení epitelů podle počtu vrstev
1.
jednovrstevný (bezobratlí) – jediný typ kromě ploutvenek, obratlovci –
všechny typy
2. víceřadý
– jádra zdánlivě ve více vrstvách, všechny buňky však dosedají na bazální
membránu (výživa). Trachea savců. Víceřadý řasinkový.
3.
vícevrstevný:
krycí
(pokožka obratlovců) - rohovatějící
výstelkový
(ústa, vagína) – nerohovatějící
4. přechodný
(uretra, moč. měchýř), podle novějších údajů jde o víceřadý epitel. Změna počtu
buněk podle stavu, ve vrchních vrstvách se nacházejí volné buňky (močový měchýř
savců).
4.2. Dělení epitelů podle tvaru buněk (URL 9)
1. kubické (žlázy, vývody)
2. cylindrické (trávicí orgány bezobratlých i
obratlovců)
3. dlaždicové:
a) jednovrstevné (pulci
obojživelníků, kopinatec)
b) vícevrstevné (epidermis u Vertebrata)
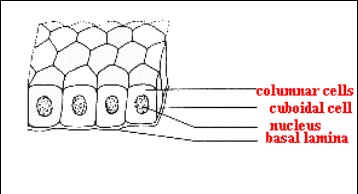

Jednoduchý kubický
epitel (URL 9)
Jednoduchý cylindrický epitel
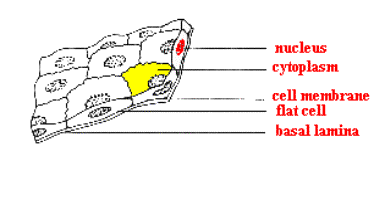
Jednoduchý
dlaždicový epitel Vícevrstvý epitel
Jednovrstevný, krycí,
žláznatý, kutikulární epitel žížaly.
Mezi epidermálními buňkami
jsou hojné váčkovité žlázky (endoepitelové) s granulemi sekretu
v cytoplazmě. Pod epitelem je silná vrstva okružní svaloviny (hladké). VP
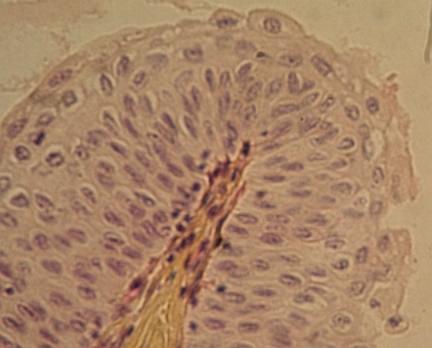
Jednovrstevný kubický epitel vystýlající žlučový kanálek
vyššího řádu v játrech. Kolem je vrstva pojiva, podlouhlá jádra patří
fibrocytům (viz dále). VP

Jednovrstevný epitel
výstelky – ependymu – míšního kanálu zárodku kuřete. Tvorba míšního moku. Jde
také
o typ neuroglií (viz dále).
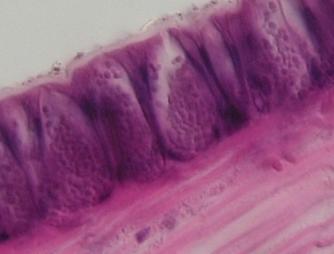
Víceřadý řasinkový epitel
průdušnice člověka. Lumen průdušnice
vlevo nahoře, vpravo dole vláknité kolagenní vazivo sliznice. VP
Přechodný epitel na vazivovém
záhybu stěny močového měchýře člověka. Podle novějších poznatků jde o epitel
víceřadý, kde každá buňka má vlastní spojení s bazální laminou pomocí
výběžku cytoplazmy. VP
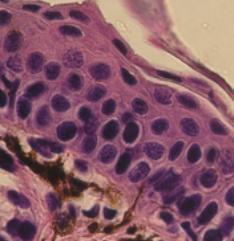
Vícevrstevný rohovatějící
(vpravo nahoře) epitel kůže žáby (Bombina sp.). Vlevo dole vrstva škáry
s pigmentovými buňkami VP
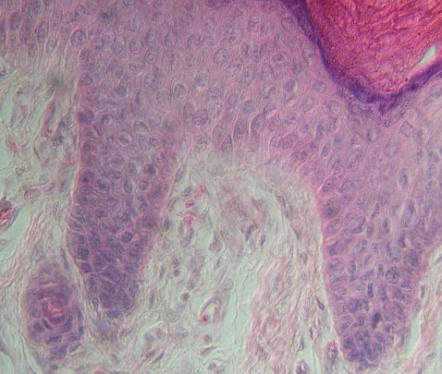
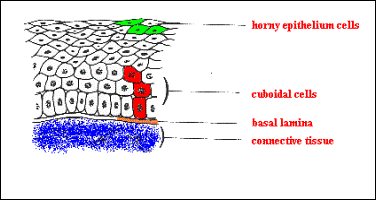
Mnohovrstevný epitel krycí,
rohovatějící v pokožce (epidermis) člověka (silná vrstva fialových buněk
v horní části obrázku) Směrem zdola se nacházejí vrstvy (stratum):
s. basale (cylindricum) dolní dělivá vrstva, regenerace
s. spinosum několik vrstev dělivých buněk. Obě tyto vrstvy tvoří tzv.
s. Malphighii.
s. granulosum – objevují se zrna rohoviny v cytoplasmě, buňky se
již nedělí,
s. lucidum – hyalinní vrstva buněk, na ploskách chodidel a dlaních
primátů, zde ano – modrý proužek vpravo nahoře
s. corneum – odlupující se vrstva buněk proměněných v šupinky
rohoviny (plné keratinu)
Těsně pod epidermis je
vrstvička řídkého vláknitého pojiva (viz dále) včetně cév, které do pokožky
nevstupují.
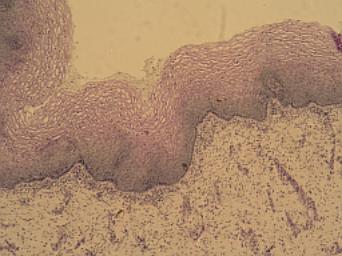
Vícevrstevný nerohovatějící
epitel labia minor ženy. VP
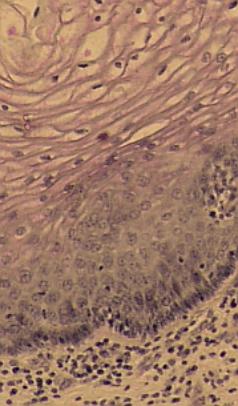
Totéž při větším zvětšení.
Zcela dole vrstva pojiva, pak stratum basale a další vrstvy epitelu. VP
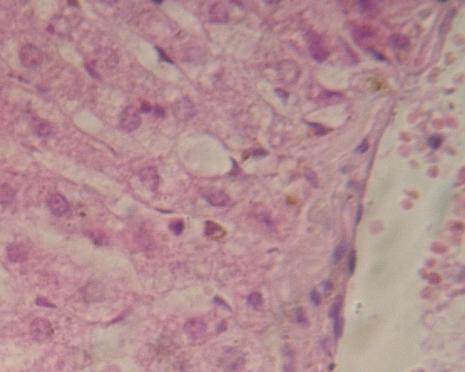
Netypický, tzv. trámčitý
epitel hepatocytů v jaterních lalůčcích. Hepatocyty tvoří prostorové
útvary obtékané krví v sinusoidech, které jsou vystlány jiným typem
epitelu – endotelem (plochá jádra v dutinách. Vpravo endotel vystýlá i
cévu, v níž jsou viditelné erytrocyty (srovnej velikost buněk – hepatocyty
jsou polyploidní, erytrocyty patří k nejmenším buňkám a u savců nemají
jádro) VP
![]()
4.3. Dělení epitelů podle polárního rozlišení
1. bičíkové - výstelka trávicího
vaku láčkovců
(jinak vzácné)
2. řasinkové - kinocilie - aktivní
pohyb:
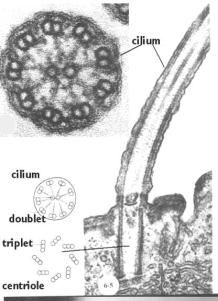 kinocilie (URL 18)
kinocilie (URL 18)
a) na povrchu těla - slouží k pohybu -
ploštěnky, pásnice
b) na povrchu
těla - k přihánění potravy - vířníci, larvy bezobratlých (trochophora)
c) ve střevech
bezobratlých - posun potravy - ploštěnky, kroužkovci, měkkýši
d) v dýchacích cestách vyšších obratlovců - posun hlenu (trachea, bronchi, bronchioli)
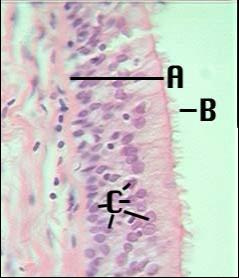
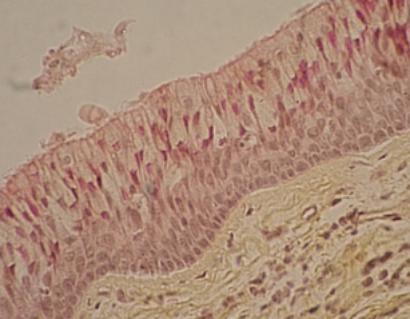
Epitel dýchacích cest. A –
vrstva epitelu (všechny buňky vycházejí z bazální membrány, ale jen
některé dosahují až k povrchu. C – jádra buněk. B – cilia. Jejich funkcí je transportovat
polutanty dýchacích cest zpět k ústům. (URL 11)
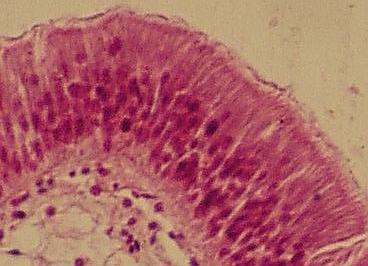
Jednovrstevný
(vlivem řezu zdánlivě vícevrstevný) cylindrický řasinkový epitel střeva škeble
(Anodonta sp.)
Stereocilie
- bez pohybu:
e) na povrchu těla mihulí - udržuje sliz

Řasinkový sloupcový epitel (URL 9)
3. mikroklky (kartáčový, žíhaný, lem) příbuzné
nepohyblivým řasinkám (vyztužené mikrofilamenty)
-
ve střevech v
resorbčním epitelu - zvýšení povrchu (záhyby, klky, mikroklky)
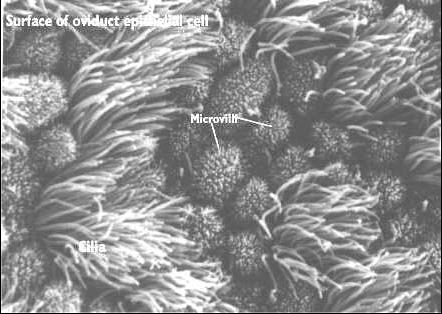
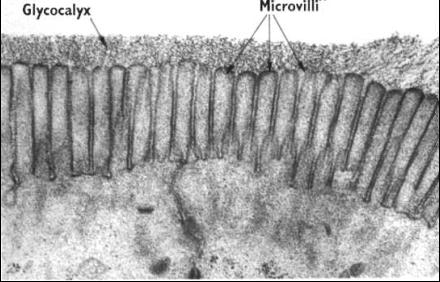
Mikroklky (microvilli) a
kinocilie (cilia) vejcovodu (URL 18)
4. límcové (URL 14) (choanocyty)
- trávicí dutina hub, přihánění a trávení potravy
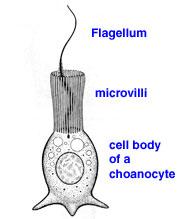 Choanocyt
(URL 15)
Choanocyt
(URL 15)
5. Kutikulární (žížala, nematoda, členovci, u hmyzu:
epikutikula-vosky, parafiny, tenká barevná (kutikulin) bílkovina, exokutikula:
silná, žlutá až černohnědá chitin + skleroproteiny, endokutikula - chitin,
nejsilnější, pružná, otvory – sekrety,odpadní látky

Kutikulární
epitel škrkavky (Ascaris megalocephala). Pod ním syncytium epidermálních buněk,
jejichž jádra jsou ponořena hluboko pod povrch těla
6. epitely nahé bez povrchových struktur
![]()
4.4.
Dělení
epitelů podle funkce
4.4.1. krycí - ochranná vrstva, drobné buňky
a) jednovrstevné: kutikulární (rak,
žížala, hmyz)
řasinkový
(Anodonta)
b) vícevrstevné: (epidermis obratlovce)
4.4.2. výstelkové - chrání vnitřek tělesných orgánů (dutina ústní,
vagína-mnohovrstev-
ný, peritoneum - mezotel - 1vrstevný, seróza)
4.4.3. Žlázové
epitely
(URL 12) - velké buňky hruškovitého nebo pohárkovitého tvaru
sekrety - užitek v dutině (sliny, trávicí
šťávy)
inkrety - hormony
exkrety - moč, pot
Žlázový
epitel (URL 9)
Žlázy podle sekrece (extruze)
-
apokrynní: výměšek odchází s částí cytoplazmy apikálního pólu
buňky (pulsace buňky podle sekrečního cyklu - mléčná žláza),
-
mezokrynní: (ekkrynní) - drobnými kanálky:
a) aktivním
transportem - (sekrece žaludeční šťávy tzv. krycích buněk)
b) plazmatická
membrána splývá s membránou sekrečního váčku a tak se sekret dostává ven (potní
žlázy, ostatní buňky žaludku)
-
holokrynní: výměšek se
hromadí v celé buňce a ta se časem rozpadá (což umožní výměšku dostat se mimo buňku (tuková tkáň, mazové žlázy kožní), buňky se
nahrazují z buněk při bazální membráně nebo z nediferencovaných fibrocytů.
Žlázy endoepitelové - (skupina buněk
v epitelu),
exoepitelové - vchlípení v
době embryonálního vývoje
Žlázy podle tvaru sekreční části:
1. alveolární
2. tubulózní
3. tubuloalveolární
Od každého typu: jednoduché, rozvětvené a složené.
Jedná se o vícebuněčné žlázy, vznikající vchlípením
epitelu (stejně jako endokrinní žlázy). Na rozdíl od nich je u nich zachováno
spojení s povrchem epitelu. Mohou nabývat různých tvarů
(URL 16):



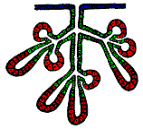
Vyvíjejí
se z krycího epitelu (modré
buňky). Tvoří jednoduché nebo rozvětvené trubičky, na jejichž koncích se
nacházejí sekreční části (červené),
které
buď zachovávají svůj tubulózní tvar nebo se rozšiřují a tvoří alveolární žlázy.
Výměsky:
plyny
(plynový měchýř ryb)
kapalina
- mucinózní (slizovité)
- serózní (tekuté)
pevná látka - exkreční
konkrementy - kutikula (schránky měkkýšů)
Žlázy
-
jednobuněčné - pohárkové buňky, pokožka a opasek
žížaly, vápenité schránky měkkýšů - v epitelu
-
mnohobuněčné - ty ostatní, endo- a exoepitelové
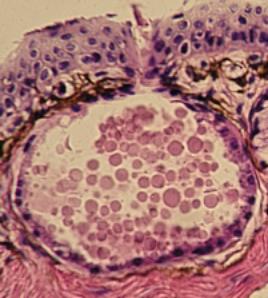
Exoepitelová, alveolární mnohobuněčná žláza
v kůži žáby. Kolem vláknité kolagenní pojivo s četnými pigmentovými
buňkami. VP
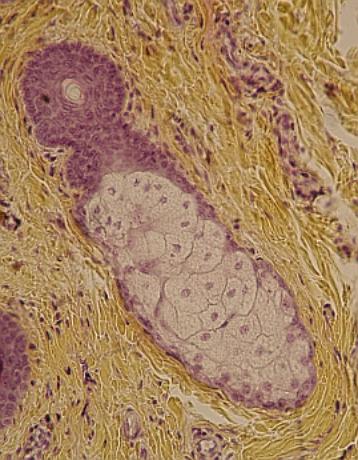
Mazová
žláza u vlasového váčku v kůži víčka člověka. Alveolární mnohobuněčný typ
s holokrinní sekrecí mazu. VP
![]()
4.4.4. Resorbční
epitely – trávicí (enterocyty),
dýchací (pneumocyty), vylučovací (podocyty)
 enterocyt (URL 17)
enterocyt (URL 17)
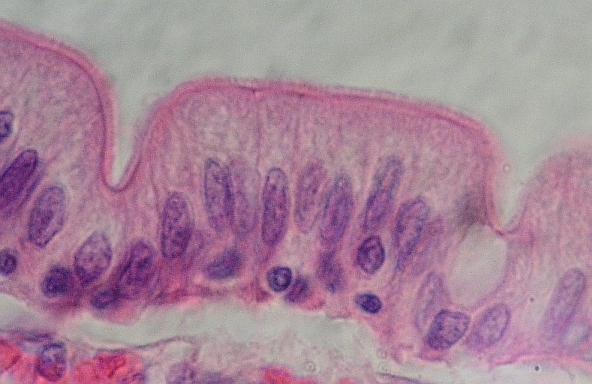
Jednovrstevný
cylindrický resorbční i žláznatý epitel
- enterocyty kryjící klky tenkého střeva člověka s tzv. žíhaným
lemem nahoře (mikroklky viditelné jen EM, pozor, neplést s klky!), vpravo
jednobuněčná (endoepitelová) žláza – pohárková buňka, vylučuje hlen.
4.4.5. Smyslové
buňky
a) primární (oční sítnice, čich, sluch) – mají
vlastní vodivé výběžky.
b) sekundární (chuť, statika, hmat) – jsou
inervovány, dendrity dostředivých neuronů
c) volná nervová zakončení (teplo, chlad,bolest) –
inervace v podstatě nesmyslových buněk epidermis).
4.4.6. Zárodečné epitely - vyživují pohlavní buňky (zrající vajíčka,
spermie), tvoří folikuly, Sertoliho buňky (obratlovci),a Versonovy buňky (pavouci, hmyz) - vyživují spermie
4.4.7. Epitely svalové - láčkovci
(Nematoda hypodermální syncytium)
4.4.8. Epitely pigmentové - pigmentová
vrstva v sítnici oka
4.5. Ontogeneze epitelů
- ektoblast - krycí epitely (potní, mazová a
mléčná žláza, dutina ústní, vagína, konečník)
- mezoblast -
výstelkové epitely druhotné dutiny tělní
-
entoblast - rezorbční epitely střev, plic, žaber, trávicí žlázy
- mezenchym -
výstelka cév
Žlázy vznikají z
toho zárodečného listu, z jehož epitelu jsou odvozené
![]()
5. POJIVA
Jde o tkáně prostorové, které obalují a spojují
orgány, vyztužují tělo, tvoří oporu a kostru v podobě různě pevných sítí až
kostí. Na rozdíl od epitelů, které neobsahují skoro žádnou mezibuněčnou hmotu,
bývají shluky buněk pojivových tkání zakotveny ve velkém množství mezibuněčné
hmoty. Tato mezibuněčná hmota je vylučována buňkami a tvoří ji vlákna bílkovin,
nacházející se v amorfní směsi proteoglykanů. Mezi ně řadíme i tělní
tekutiny, které rovněž spojují orgány, rozvádějí výživu, hormony a obranné
látky, odvádějí zplodiny metabolismu a případně rozvádějí kyslík a odvádějí CO2.
Většina pojiv
vzniká z mezoblastu (entomezenchym), vzácně z ektomezenchymu.
Mezenchym vzniká z mezoblastu.
5.1.
Struktura pojiv
-
Buňky: pevné (fixní) a
volné
-
Mezibuněčné hmoty:
vláknitá (fibrilární) a interfibrilární (to co je mezi vlákny)
5.1.1. Pevné buňky (fixní)
- větvené s bohatými cytoplazmatickými
výběžky
- oválné, polygonální, izolované
buněčnou hmotou
- syncytiální
útvary ze splynutých buněk
- (pigmentové buňky)
5.1.2. Volné buňky
- mohou se
pohybovat v tekutých mezibuněčných hmotách a mají často i schopnost fagocytózy.
Bývají amoeboidního tvaru a diferencují se někdy z buněk fixních, jindy do
pojiva vcestují např. z cévního oběhu.
Žírné buňky
(heparinocyty,
mastocyty) – nacházejí se
všude v řídkém pojivu, bývají kulovité, s malým jádrem, a obsahují
v granulích heparin (kyselý mukopolysacharid – snižuje srážlivost
krve), histamin (permeabilita kapilár, snižuje krevní tlak), serotonin
(zvyšuje krevní tlak, zesiluje peristaltiku střev).
Histiocyty
- makrofág, klasmocyt - umí fagocytovat (hojení ran, hnisavé záněty), mají
menší jádro a mnoho lyzozómů. Odlišení: vitální barvení - fagozómy s
jednotkovou membránou. Fixní a aktivované (pohyblivé) makrofágy. Vznik
z monocytu.
(Srovnej mononukleární fagocytární systém: monocyt,
histiocyt, Kupferova buňka, prachový makrofág, mikroglie, slezinné makrofágy,
osteoklasty.)
Granulocyty -
eosinofilní granulocyty - někdy i neutrofilní (bílé krvinky) - vycestovavší z
krevního oběhu, schopné pohybu, fagocytózy, obrana organismu (mikrofágy) –
nespecifická imunita (např. ve stromatu mléčné žlázy, intersticium plic… všude
v řídkém vláknitém pojivu).
Lymfocyty
- identické s krevními lymfocyty, účastní se imunitních reakcí specifické
imunity . Dávají vznik i dalšímu typu volných buněk:
-
plazmatickým buňkám - typické loukoťovitým chromatinem v jádrech,
vyrábějí protilátky specifické imunity, nemají Golgiho aparát, váčky přímo
z drsného ER.
-
též epitelovým
buňkám za určitých patologických
stavů.
5.1.3. Mezibuněčné hmoty
Vláknité hmoty - kolagenní, elastické, retikulární (podle převládajícího typu
bílkovin.)
Funkce: zpevňovací (zpevnění vlákna), zvláštní
skupinu tvoří krevní vlákna (fibrin).
a) Vlákna kolagenní – vláknitá bílkovina s vysokým % hydroxiprolinu, velmi pevná, málo
protažení schopná, varem rozpustná, po ochlazení klihovatí. Každé vlákno
z protofibril tropokolagenu, 10-40 nm tlustých. Vlákna tvoří zvlněné
pentlice o průměru
1-2 μm, (dvojlom - jemné žíhání 1/4 délky).
Nacházejí se ve velkých množstvích v těle většiny
živočichů. Chybějí u Protozoí a velmi málo se vyskytují u
Arthropod.
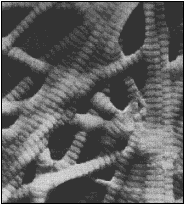 Kolagen I.
- tuhá pojiva vláknitá, plsťovitá,provazcovitá (šlachy, kostní tkáň, dentin v
zubech).
Kolagen I.
- tuhá pojiva vláknitá, plsťovitá,provazcovitá (šlachy, kostní tkáň, dentin v
zubech).
Kolagen II.
- chrupavčité tkáně, struna hřbetní, sklivce.
Kolagen III.
- střední vrstva cév, tepen a žil, škára
Kolagen IV.
- bazální membrána (lamina densa)
Kolagen V.
- pojivové tkáně kromě chrupavek, váže se na kolagen I.
Všechny (I.-V.) kolageny = 25% všech bílkovin
u obratlovců.
kolagen (URL
19)
b) Vlákna
elastická
(hlavní součástí je bílkovina elastin). Jsou odolná vůči varu, kyselinám i
zásadám. Mají žlutavou barvu a hedvábný
lesk. Pod elektronovým mikroskopem vidíme jemné podélné žíhání.
Jednolomné, zřetězené globule elastinu, tahem protažitelná ve vlákna dvojlomná,
150% roztažnost, síla 1-10 μm, síťovitě
rozvětvená v kolagenním vazivu (valin, glycin, prolin, alanin). (Podle novějších studií podstatně
komplikovanější složení.)
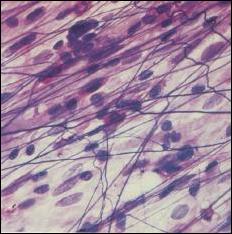 Řídká vláknitá pojivová tkáň (Loose Areolar
Connective Tissue) (URL
20)
Řídká vláknitá pojivová tkáň (Loose Areolar
Connective Tissue) (URL
20)
c) Retikulární vlákna jsou strukturou podobná kolagenním, retikulin
(heteroprotein s vysokým obsahem glycidických složek).
Vlákna se skládají z protofibril, avšak plsťovitě a
síťovitě propletených.
Interfibrilární hmoty: mukopolysacharidy a bílkoviny + nerozpustné
minerální soli
-
kyselina hyaluronová –
slizovitá
-
kyselina mukoitinsírová
– rosolovitá
-
kyselina
chondroitinsírová – pevná
-
minerální soli - v kostech a ve stáří v chrupavkách
-
tělní tekutiny - zvláštní typ mezivláknových hmot
5.2. Druhy pojiv
Podle rozmanitého
zastoupení výše uvedených prvků najdeme v literatuře následující druhy
pojiv:
Mezoglea
Zárodečný
mezenchym
Rosolovité pojivo
Retikulární
(síťovité) pojivo
Řídké vláknité
pojivo
Tukové pojivo
Tuhé vláknité pojivo – vazivo
Pojivo buněčné
Pojiva chrupavková
Kost
Zubní
tkáň
Pojiva trofická
(tělní tekutiny)
5.3. Mezoglea (u
Diblastica)
Nejprimitivnější pojivo z hlediska fylogeneze.
U hub a žahavců - oporná funkce. (URL
27, URL
28)
Základní hmota – činnost ektodermu, entodermu a
vycestovaných buněk.
Spongioblasty (amoeboidní tvar) v mezoglei hub (spongin)
Skleroblasty
v mezoglei hub a žahavců, produkujících křemité a vápenité skelety.
Archeocyty
- nediferencované mezogleální buňky - buňky pohlavní, spongioblasty,
skleroblasty.
5.4. Zárodečný (embryonální) mezenchym
= pojivová
tkáň v zárodcích. Z ní diferencují všechna ostatní
pojiva (kromě
chordy), hladká svalovina, chrupavka, kost (i buňky
krevní, krev.
cévy). Vzniká z buněčného materiálu mezodermálního původu.
Buňky hvězdicovité,
tenké, rozvětvené plasmodesmy = prostorová síť spojených plazmodesem. V
ní i buňky volné, amoeboidní.
Základní hmota
tekutá
s příměsí mucinózních látek (tkáňový mok). Fibrilární hmoty vzácně
- retikulární fibrily. U některých
i v dospělosti
- parenchym (u ploštěnek, částečně i pijavek i jiných kroužkovců).
Retikulární fibrily v amorfní substanci. Množství
roste s postupnou diferenciací pojiva.
Z něj: všechna vaziva (kromě chordy), kost,
chrupavka, krev, cévy, cévy krevní i lymfatické
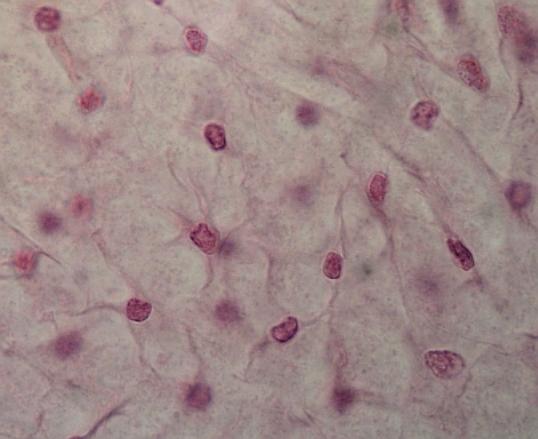
Embryonální
mesenchym lidského zárodku
Hvězdicovité
pluripotentní buňky, spojené cytoplazmatickými výběžky, začínají tvořit vlákna.
Někdy jsou buňky vřetenovité. Mezibuněčná hmota vodnatá, nezřetelná. (Tu a tam
pohyblivé buňky.) Srovnej též (URL
22) Slide
1
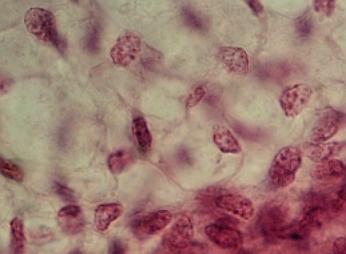
Zmenšením mezibuněčných prostor (vpravo dole) a následnou diferenciací buněk vzniká z embryonálního mezenchymu tkáň. VP
5.5.
Rosolovité pojivo
Podobné
zárodečnému mezenchymu, volné buňky méně početné a mezibuněčná hmota je rosolovitá
s vysokým obsahem mucinu (mukoitinsírová kyselina a bílkovina). Přítomny v
menším množství retikulární nebo kolagenní vlákna. Vyskytuje se v
zárodcích, v okolí orgánových základů a v pupečním provazci savců
(pupečník - funiculus umbilicalis, Whartonův rosol (URL 21) = embryonální charakter)
hvězdicovité buňky – fibroblasty spolu s makrofágy -bloudivé buňky
mezibuněčná hmota rosolovitá s retikulárními vlákny
Stěna cév pupečníku = ze snopců
okružní, podélné a spirální hladké svaloviny, oddělených vrstvičkami
rosolovitého vaziva.
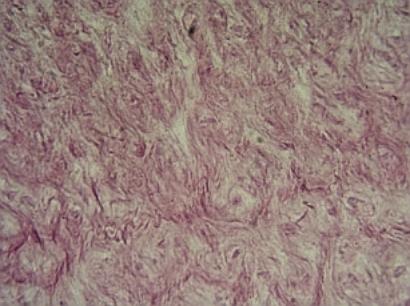
Whartonův rosol
v pupečníku plodu člověka. Skladba:
Hvězdicovité buňky, fibroblasty, makrofágy,
mezibuněčná hmota rosolovitá, mnoho mucinu, tvorba nemnoha kolagenních i
retikulárních vláken.
Srovnej též (URL 21).
5.6. Retikulární (síťovité) pojivo
Podobné zárodečnému a rosolovitému pojivu. Prostorová syncytiální síť z hvězdicových buněk (retikulární buňky) produkujících retikulinová vlákna. Ta se nacházejí z části v buňkách, z části tvoří mezibuněčnou vláknitou hmotu. Mezibuněčná hmota je jinak tekutá s početnými volnými buňkami.
Výskyt:
orgánové základy embryí
1. schopnost další
diferenciace
2. v dospělosti tvoří obalové
sítě kolem jaterních trámců a ledvinných kanálků
3. samostatné orgány
- soustava retikuloendoteliální (RES):
slezina, endotel krevních cév + lymfatické uzliny /mandle/, červená kostní
dřeň). Zde bývá v pojivu mnoho volných buněk,
především lymfocytů (vznik z retikulárních buněk přes lymfoblasty).
(Mononukleární fagocytární systém - diferencované makrofágy mimo krevní řečiště: histiocyty v řídkých vláknitých pojivech, slezinné makrofágy, Kupferovy
buňky v jaterních sinusoidách, alveolární, tzv. prachové makrofágy v plicních sklípcích, mikroglie v nervové tkáni, asi též chondroklasty, což je totéž co osteoklasty – obří makrofágy likvidující chrupavku
a kost, vzniklé splynutím několika monocytů).
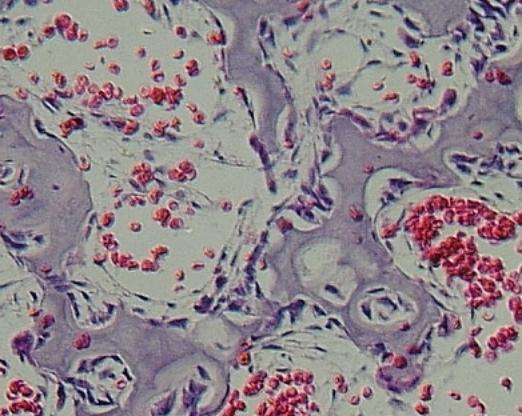
Retikulární
pojivo při primární enchondrální osifikaci ulny lidského plodu.
Tvoří síť
buněk v prostorách mezi prvními kostními trámci (fialové), z nichž
některé se mění v osteoblasty a přisedají ke kosti, jiné tvoří budoucí
buňky krevní, z dalších vzniká endotel kapilár, které odvádějí
diferencované buňky krevní – červené krvinky. VP
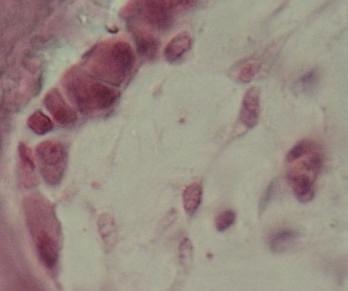
Retikulární
buňky (vpravo) se mění v osteoblasty (vlevo),
které
přisedají k trámečku primární kosti. VP
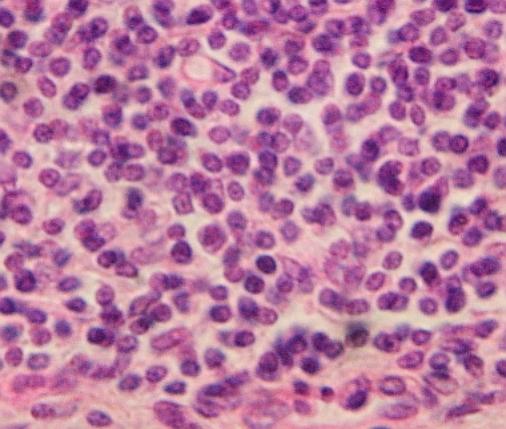
Retikulární
pojivo tvoří též lymfatické uzliny, na obr. snímek z tzv. Peyerova plaku
v tenkém střevě člověka. Prostor mezi retikulárními buňkami (větší, méně
zřetelná jádra) je zcela vyplněn lymfocyty. VP
Pro další detaily retikulárního pojiva viz též URL 22:
Slide
4: Hvězdicovité buňky, schopné
diferenciace, spojené plasmatickými
výběžky, retikulární vlákna jakoby v cytoplazmě. Základ červené kostní
dřeně a lymfatických orgánů. Tam pak je v mezibuněčných prostorách mnoho
lymfocytů, případně makrofágy. Zde detail lymfatické uzliny.
Slide
2: Struktura stříbřené lymfatické tkáně.
Slide 3: Hvězdicovité
retikulární buňky (světlé) tvoří jemnou síť výběžků doplněných retikulinovými vlákny
(stříbřením zviditelněná).V mezibuněčných prostorách lymfocyty. Preparát lymfatické uzliny.
![]()
5.7. Řídké
vláknité pojivo
Nejrozšířenější výplňové pojivo navíc i s
funkcí obrannou a vyživovací, regulace vody v těle, zásobárna
tuků a glykogenu).
Oporné soustavy (stroma) a obaly téměř všech tělních
orgánů, ovšem jako součást hustého vláknitého pojiva, které je pevné a jehož
buňky potřebují výživu, energii a ochranu.. Proniká dovnitř tkání, o jejichž
buňky pečuje.
Najdeme je např. v
horní vrstvě škáry (pars papilaris cori), kůže a v tunica propia
sliznic. Je mezi svalovými vlákny, vyplňuje prostory mezi lalůčky žláz atp.
Obsahuje:
Fibrilární hmoty - vlákna kolagenní a elastinová
plsťovitě uspořádána.
Interfibrilární hmota - slizovitá až řídká, tekutá (tkáňový mok).
Viskozita podle obsahu mucinózních látek. (Hromadění tkáňového moku - otoky).
Složení:
1.
fibrocyty
2.
elastické fibrily
3.
kolagenní fibrily
4.
tkáňový mok
5.
plazmatické buňky
6.
histiocyty (klasmocyty)
7.
žírné buňky
8.
granulocyty (neutrofilní, eosinofilní)
5. lymfocyty
5.7.1. Fixní buňky řídkého vláknitého
pojiva
1) Fibroblasty = fibrocyty (desmocyty) a dále
a) buňky hvězdicového typu
b) buňky tukové
c) buňky pigmentové (přicestují)
a) Marchandovy buňky (kombiální buňky) -jsou
důležité pro hojení ran. Diferencují všechny typy buněk (zárodečné
mezenchymální buňky) a rozrůstání pletiva - dosud nediferencovaného
b) Pigmentové buňky(chromatofory) se dělí na:
melanofory (černý melanin), iridiocyty (stříbřité quaninové krystaly),
xantofory (žlutá barviva -karoteny), erytrofory (červená barviva). Některým
živočichům umožňují barvoměnu.
c)Tukové buňky (viz dále).
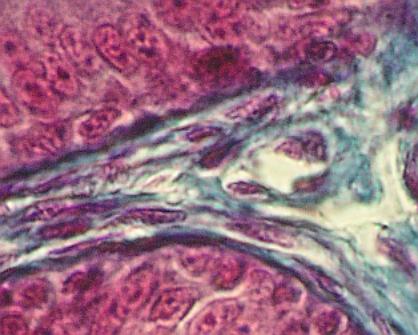
Řídké vláknité
pojivo v lamina propria mucosae sliznice tenkého střeva skokana (Rana sp.).
Uprostřed fialová protáhlá jádra fibrocytů, modře zbarvená kolagenní vlákna.
Nahoře a dole enterocyty klku, jednovrstevný epitel resorbční. Povšimněte si
tmavé linie bazální laminy na rozhraní epitelu a pojiva. Uprostřed poněkud
vpravo je příčný řez lymfatickou cévou.
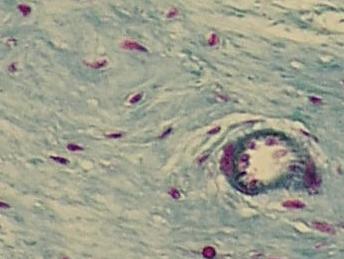
Řídké
vláknité pojivo podslizničního vaziva žaludku skokana Rana sp. Jádra fibrocytů
fialová, kolagenní vlákna modrá. Vpravo krevní céva, výstelka – endotel (modrá
kulatá menší jádra)
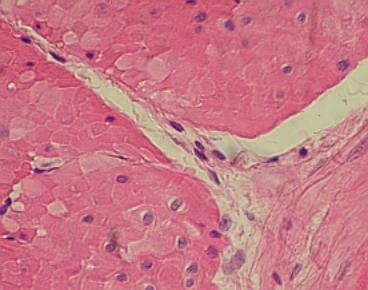
Řídké
vláknité pojivo (světlé linie uprostřed s typickými jádry fibrocytů a
převážně kolagenními fibrilami) propojuje úseky hladké svaloviny v tunica
muscularis žaludku kočky (Felis sp)
Pro další srovnání viz též URL 22:
Slide
21 Diferenciací
EM vznikají fibroblasty a z nich fibrocyty – fixní buňky řídkého vláknitého
pojiva. Do mezibuněčné hmoty produkují kolagenní a elastická vlákna (někdy
primárně retikulární - v mládí)
Slide
22 Jiný fibroblast mezi snopci kolagenních
vláken (růžově zbarvené). Typické je podlouhlé jádro.
Slide
8 Sítivo vláken v řídkém vláknitém
pojivu. Silnější přímá vlákna – kolagen (žlutá), tenká vlnitá – elastin (bílá).
Většina jader náleží fibroblastům. (URL 22).
Slide 6 ŘVP spojuje tkáně, zde mnohovrstevný epitel. Umožňuje přístup cévám, nervům a zajišťuje ochranu. Hospodaření s vodou – vznik lymfy, otoky. (URL 22)
Slide 7 ŘVP upevňuje cylindrický epitel střeva (enterocyty). Tvoří strukturu klků. V takovém případě má též mnoho lymfocytů. (URL 22)
Slide
5 ŘVP (modře) kolem tubulů
ledvinných kanálků obsahuje i retikulární vlákna (stříbřena, černá), která však
byla vyloučena fibroblasty. Pouze v kostní dření, slezině a lymfatických
uzlinách jsou retikulární vlákna produkována retikulárními buńkami. Kolagenní
vlákna jsou jemná, těsně kolem tubulů. (URL
22)
Slide 23 Několik fibroblastů mezi snopci ŘVP. (URL 22)
Slide 24 EM snímek fibroblastu sekretujícího tropokolagen (vně buňky). Silně vyvinuté EPR s cisternami plnými prekursoru bílkoviny. (URL 22)
ŘVP upevňuje cylindrický
epitel střeva (enterocyty). Tvoří strukturu klků. V takovém případě má též
mnoho lymfocytů. Umožňuje přístup cév krevních, mízních, inervaci, obranu.
Červeně zbarvené jsou erytrocyty v kapilárách o stejném průsvitu jako krvinky.
VP
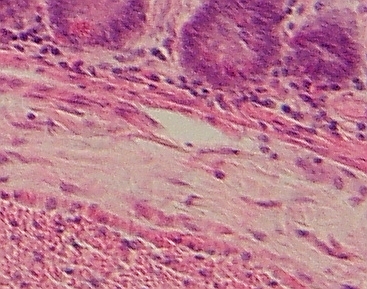
Řídké
vláknité pojivo tvoří vrstvu podslizničního vaziva (široký světlejší pruh
napříč obrázkem), která spojuje sliznici (nahoře) s vrstvou svaloviny
(dole). Zřetelná jsou podlouhlá jádra fibrocytů. ŘVP proniká volně do
svaloviny, kde tvoří epimysium svalových buněk i nahoru do sliznice, kde
podpírá slizniční vychlípeniny – řasy a klky.
![]()
5.7.2. Volné buňky
řídkého vláknitého pojiva
1)
Histiocyty,
klasmocyty, makrofágy (synonyma) -
amoeboidní tvar, malé jádro, zrnitá cytoplazma. Fagocytují velké částice.
Hojení ran, záněty (granula - rozpadlé fagozómy)
2) Žírné buňky (heparinocyty, mastocyty) – ploché,
oválné, protisrážlivá látka (heparin),
zachycují škodlivé látky, fagocytózy neschopné, bazofilní zrna v cytoplazmě (heparin,
histamin, serotonin)
|
Žírné buňky v ŘVP plné sekrečních váčků s prekursory
histaminu a heparinu. Imunitní reakce, alergie. (URL
22) |
Žírné buňky. Tmavá barva granulí, které se dostaly mimo buňky
histologickým zpracováním preparátu. Jádra světle modrá. (URL
22) |
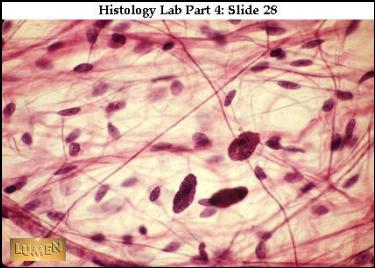
ŘVP s žírnými buňkami. Většina
ostatních jader náleží fibrocytům. (URL
22)
|
EM snímek žírné buňky. (URL
22) |
EM lidských žírných buněk ukazující různou strukturu sekrečních váčků. |
3) Plazmatické buňky – vejčitého tvaru, schopnost pomalého amoeboidního pohybu.
V jádrech chromatin soustředěn.
"loukotovitě", silně bazofilní, cytoplazma
ale homogenní. Hlavní funkce – tvorba protilátek. Hlavně v mízních uzlinách,
blanách obalujících orgány, ve střevní sliznici a při chorobných stavech i v
krevním oběhu.
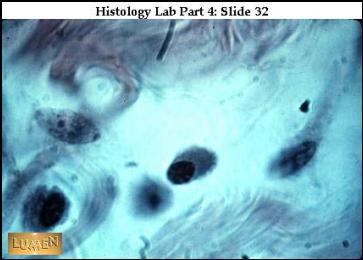
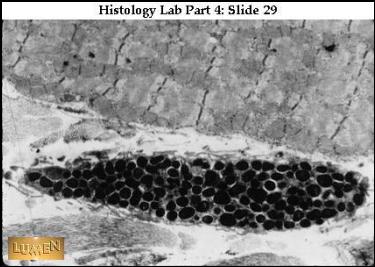
|
Plazmatické
buňky – vznik z B lymfocytů, výroba protilátek. Bazofilní
cytoplazma, excentricky situované jádro s loukoťovitě umístěnými bloky
heterochromatinu. Světlý okrsek vlevo od jádra je negativ Golgiho aparátu.
Mnoho růžově zbarvených vláken kolagenu v mezibuněčném prostoru. |
Jiné plazmatické buňky. Velké jádro vpravo – fibrocyt. (URL 22) |
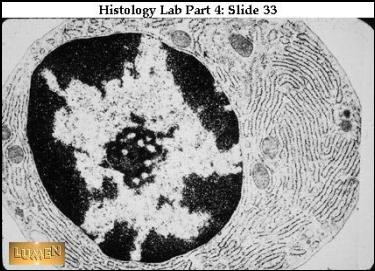
Plasmatická buňka v EM – nukleolus a loukoťovitě umístěný chromatin. Mnoho cisteren R-EPR v cytoplasmě (protilátky).
4) Ostatní volné buňky
– v řídkých pojivech jsou buněčné elementy krve a mízy – monocyty,
lymfocyty, granulocyty (eosinofilní, neutrofilní).
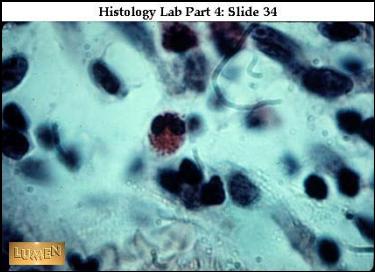
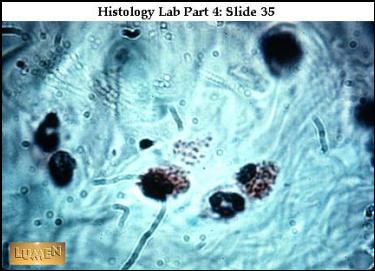
|
Eosinofilní granulocyt v mezibuněčném prostoru ŘVP –
mikrofág. (URL
22) |
„Putující“
eosinofilní a neutrofilní granulocyty v ŘVP
|
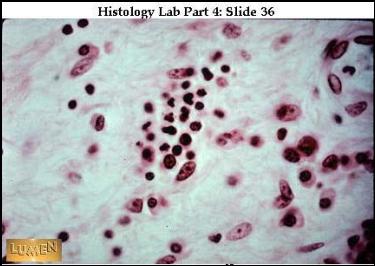
Různé buňky v řídkém vláknitém pojivu. Jádra bez cytoplasmy – asi
lymfocyty. V centru neutrofily, velká světlá roztroušená jádra – fibroblasty.
Vpravo v rohu plasmatické buňky. Vpravo nahoře céva s endotelem a neutrofilní
granulocytem (URL
22)

EM snímek volného
lymfocytu Jádro a malý okrsek cytoplasmy.
Prekurzory
plazmatických buněk. V lymfatických orgánech i jiné druhy.
Funkce řídkého vláknitého pojiva: podpůrná, látková výměna, voda, proteiny v mezibuněčné hmotě,
elektrolyty – vliv na skladbu krevní plazmy, obrana organismu (místní záněty),
hojení ran, regenerace (fibroblasty), korekce vitamínu C (vliv výživy) –
nedostatek vaziva, netvoří se kolagen.
![]()
5.8. Tukové
pojivo
Modifikace řídkého vláknitého pojiva, kde se
embryonální mezenchym diferencoval v lipoblasty.
Fixní buňky
– (embryonální lipoblasty, z nich adipocyty) kulovité, objemné tukové
buňky (URL 30),
které se navzájem dotýkají. V mezerách mezenchymová tkáň (růst tuk.
pojiva) + fibrilární mezibuněčná hmota. Cytoplazma s jádrem je postupně
zatlačena k povrchu buněk vakuolami tuku. Mezi buňkami se nachází velice málo
mezibuněčné hmoty a vláken. Pouze malá část na okraji buňky obsahuje cytoplazmu
a jádro.
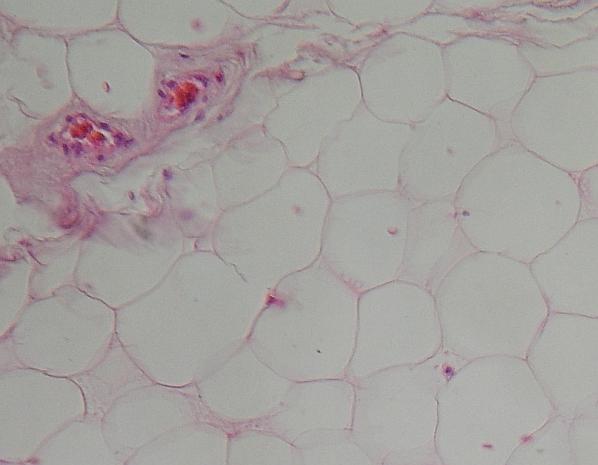
Tukové
pojivo v epineuriu člověka. Vrstvy adipocytů jsou protkány ostrůvky řídkého
vláknitého pojiva, které umožňuje vaskularizaci (přívod cév) – vlevo nahoře.
Červené buňky jsou erytrocyty.. Z nediferencovaných buněk této oblasti
může snad TP narůstat. VP
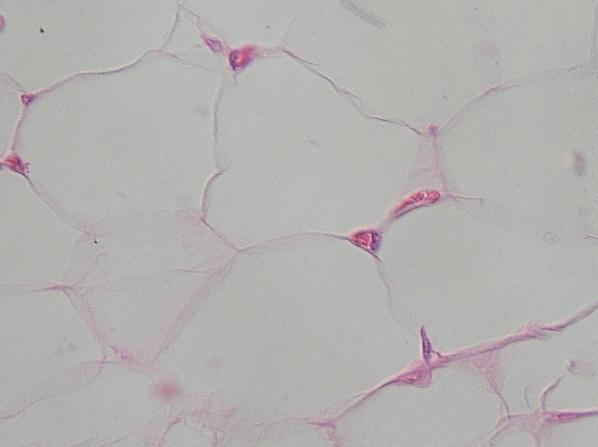
Detailní
snímek TP (z epineuria člověka) ukazuje, že každý adipocyt má kontakt
s nejjemnějšími kapilárami. Mezery mezi buňkami jsou vyplněny vlákny a
volnými buňkami typu řídkého vláknitého pojiva, z něhož je TP odvozeno. VP
Význam:
hlavní zásobárna chemicky vázané energie, tepelná izolace, mechanická
ochrana vnitřních orgánů.
U obratlovců: pod kůží, na povrchu orgánů.
Existují dva typy TP - bílá tuková tkáň
(typická, tzv. univakuolární) a hnědá
tkáň (multivakuolární): červenohnědá barva, mnoho kapilár, typ buněk,
chemický ohřívač organismu (děti do 3 let, zimní spáči). Pomocí termogeninu
mění mitochondrie energii nikoli na chemickou, ale na tepelnou. Teplo se
rozvádí bohatě přítomnými cévami.
![]()
5.9. Tuhé vláknité
pojivo - vazivo
Vznikají ze zárodečného mezenchymu tam, kde jde o tah
nebo tlak. - převaha fibrilární mezibuněčné hmoty (kolageny). Volné buňky
tam téměř nejsou, stejně je málo i hmot interfibrilárních.
Fixní buňky:
drobné polygonální fibrocyty.
5.9.1.
Plsťovité
vazivo
- snopce kolagenních vláken, jednotlivě elastinová vlákna, vše je plsťovitě
propletené. Drobné fibrocyty se nacházejí ve štěrbinách mezi svazky
vláken.Tvoří spodní vrstvu kůže, tzv. škáru (pars reticulare cori).
Podílí se na stavbě cévních stěn (zacelování ran), vzdoruje tlaku (např.bělima
oka), spoluvytváří pouzdra obalující orgány (fibrózní blány, fascie).
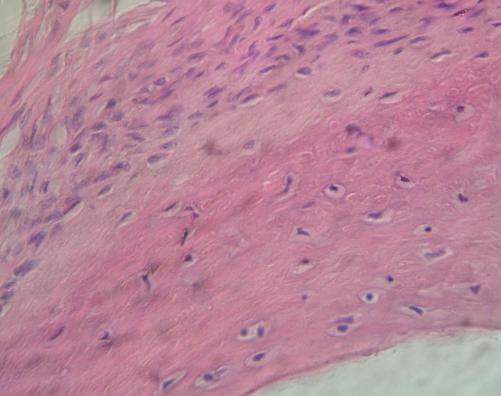
V levém horním rohu je okostice z hustého
vláknitého pojiva plsťovitého na kosti z článku prstu dítěte. Zřetelná
jsou nahuštěná jádra fibrocytů i fibroblastů, z nichž některá volně
přecházejí v osteoblasty povrchových vrstev kosti (střední část obrázku)
Kolem fibrocytů je velké množství těšně přiléhajících vláken (převážně
kolagenních). VP
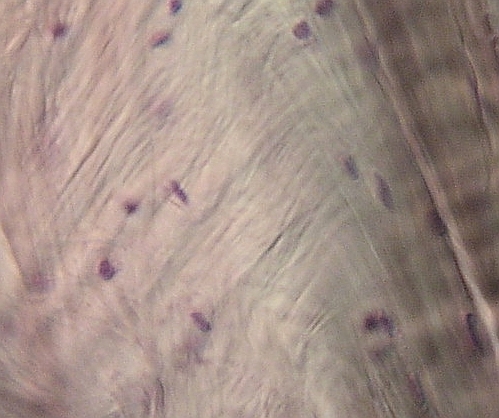
Velmi
hutné kolagenní vazivo v čéšce člověka, přechod mezi plsťovitým a
vláknitým typem. Fibrocytů je ve srovnání s mezibuněčnou hmotou málo. VP
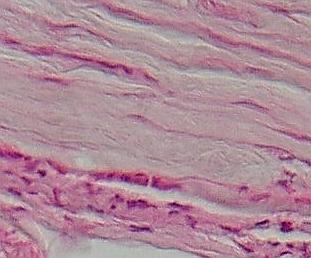
Vláknité
kolagenní pojivo epineuria zrakového nervu kočky. V horní části husté
vazivo, dole spíše řídké vláknité pojivo umožňující přísun energie (cévy) a
ochrany (volné buňky) do nervové tkáně (dole, v obrázku již nezachycena).
VP
![]()
5.9.2. Provazcovité vazivo - svazky fibril kolagenních a menší množství
elastických jsou uspořádány podélně.. Pevnost v tahu - šlachy a
fascie svalů v oblasti proměny ve šlachu. Jinak obaly svalů
(epineurium) – husté vláknité pojivo plsťovité.
Fibrocyty šlachy, zde zvané jako „křídlaté buňky“,
jsou stěsnané mezi snopci vláken s plochými výběžky cytoplazmy mezi nimi:
1) endotenonium (obal prim. svazku),
2) epitenonium (sekundární svazek)
3) peritenonium (povrch šlachy – pouzdro)
Někdy synoviální pochva, synoviální mok.
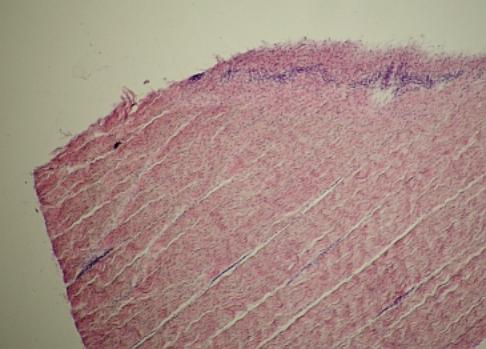
Podélný
řez šlachou – na povrchu kolagenní vazivo (peritenonium), které směrem dovnitř
rozděluje tkáň na menší úseky pronikáním řidšího pojiva (epitenonium). Existuje
ještě třetí úroveň dělení tkáně (endotenonium – kolem jednotlivých svazků
fibrocytů (zde málo zřetelné). VP

Podélná
struktura šlachy – jednotlivé úseky odděleny řídkým vláknitým pojivem
(epitenonium a endotenonium - ochrana, vaskularizace, inervace). VP
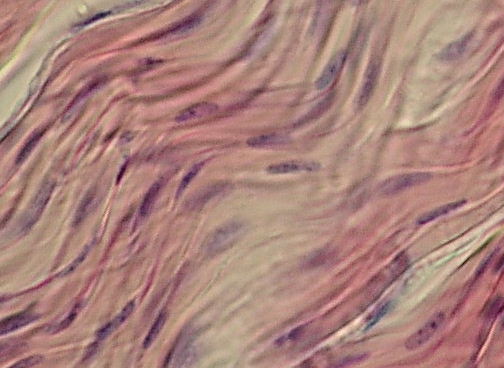
Detail
uvolněné šlachy – fibrocyty (křídlaté buňky) jsou obaleny masou podélně
uspořádaných kolagenních vláken. Při dolním a horním rohu se nacházejí jemné
světlejší vrstvičky ŘVP – endotenonia. VP
Srovnej též (URL
22)
5.9.3. Vaziva elastická - podobná jako provazcovitá, avšak s převahou
síťovitě pospojovaných elastinových vláken - pružné spoje (vazy v hlasivkách,
podélné spoje obratlů, stěny tepen).
5.10. Pojivo
buněčné – na pohled podobné
tukovému, velké dotýkající se vakovité buňky s vakuolami vyplněnými vodnatou
hmotou zajišťující tzv. buněčný turgor - pružnost a pevnost tkáně.
Mezibuněčných hmot málo.
Opora chapadel láčkovců i hlavonožců (obr níže),
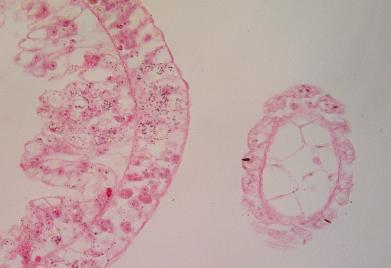
Příčný řez
nezmarem (Hydra sp.), vpravo řez chapadlem, které je uvnitř vyztuženo buněčným
pojivem
Osní orgán strunatců: chorda dorsalis - struna
hřbetní
Buněčné pojivo chordy jediné, které se diferencuje přímo z endoblastu,
(avšak nikoli vždy). Základní buňky – chordoblasty se diferencují
v chordocyty. Po celý život se nachází u kopinatců, a kruhoústých. Jinak
jen zbytky chordy mezi obratli. Pokud je v larválním výboji, bývá
nahrazeno chrupavčitou nebo kostěnou kostrou (obratel).
Struna
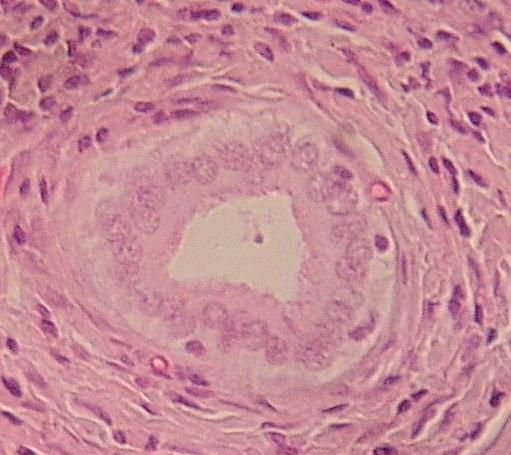
hřbetní (Chorda dorsalis) larvy čolka. Uprostřed zralé chordocyty, po stranách,
blíže k vazivovému obalu chordoblasty.
![]()
5.11. Pojiva chrupavková -
chrupavka (cartilago) - opora, pružnost, vzdor tlaku.
a) fixní buňky – chondrocyty
b) fibrilární hmoty – kolagenní i elastinové
c) interfibrilární hmota - chondrin
Diferenciace - ze zárodečného mezenchymu. Fixní
mezenchymové buňky, zakulacení, chondroblasty = zvětšení, růst,
oválné chondrocyty. Tekutá mezenchymová mezibuněčná hmota nahrazena chondrinem.
V chondrocytech - dosti tuku a glykogenu, chondrin (chondromukoid) -
skleroprotein (chondroitinsírová kys. + albuminy).
Chondrocyty v kapsulách - v pouzdrech z chondrinu (bez kolagenních vláken).
Dělením chondrocytů vznikají shluky izogenní skupiny. Okolo pouzder
chondrocytů i celých izogenetických skupin se tvoří acidofilní dvorce - teritoria
z chondrinu a husté sítě kolagenních vláken. Interteritoriální
prostor = bazofilní hmota (chondrin + menší množství elastických a
kolagenních vláken).
Chondroitinsírová A = 2x galaktozamin = (disacharid
chondrosamin)
- zbytek kys. octové, sírové a glukuronové + albuminy
= chondrin
Kolagenní vlákna - acidofilní - jsou bazická,
chondrin je silně bazofilní - kyselý.
Ochrustavice
(perichondrium) povrchový pojivový obal - na povrchu – ochrana, vstup cévy,
nervy. Chrupavka sama je necitlivá.
Růst
apozicí a za mlada intususcepcí = izogenní skupiny, které vylučují novou
mezibuněčnou hmotu.
Typy chrupavek
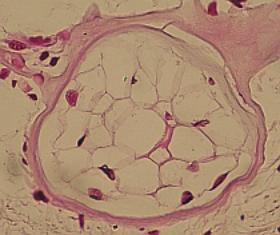
a) Pojiva chondroidní - fylogeneticky primitivní - Cyclostomata,
Plagiostomi (žraloci, rejnoci, kruhoústí, příčnoústí, částečně i u kostnatých
ryb. Málo mezibuněčných hmot, chondrocyty „bublinovitě“ vyplňují prostor
chrupavky.
Často zachovány plazmodesmy chondrocytů. Podobná je chrupavka
parenchymová u larev obojživelníků, též ucho myší.
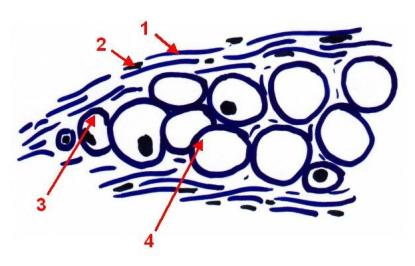
Cartilago –
chrupavka – parenchymová:
Velké
chondrocyty s malým množstvím mezibuněčné hmoty
1 - Perichondrium (vláknité pojivo)
2 - Jádra fibroblastů (f-cytů)
3 -
Chondroblasty
4 -
Chondrocyty
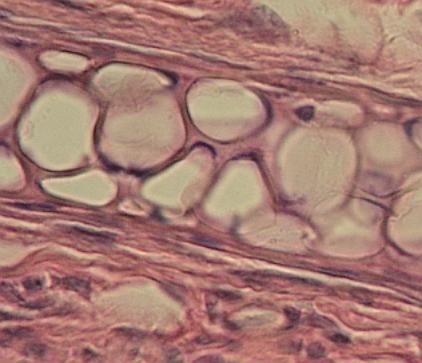
Parenchymová
chrupavka ušního boltce myši (Mus domesticus). Pás kulovitých chondrocytů uprostřed,
nahoře a dole ochrustavice, dolní vrstva silnější s příčně říznutými
cévami.cévami.VP
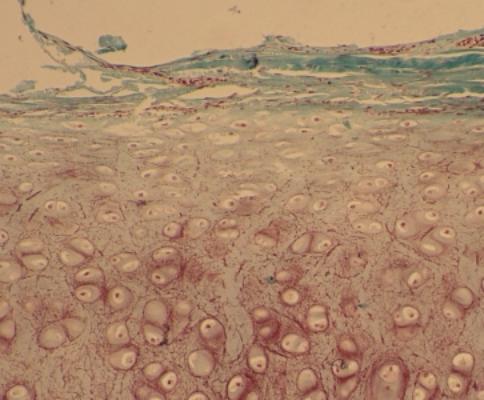
b) Chrupavka
hyalinní - průsvitná, opaleskující, lehce namodralá. Index lomu fibril
i chondrinu je stejný, proto se jeví jako homogenní Je fylogeneticky i ontogeneticky
původní, předchází vývoji ostatních typů i kostní tkáně. - Vzácně:
mnohoštětinatci, plži, hlavonožci. Celý život je u žralokovitých a
skelnošupinatých. U vyšších obratlovců: z embryonálního mezenchymu, nebo více
diferencovaného vaziva, později může být nahrazena kostí - hrtanová chrupavka,
konce žeber, některé kloubní plošky, nosní přepážka, stárnutím kalcifikuje.
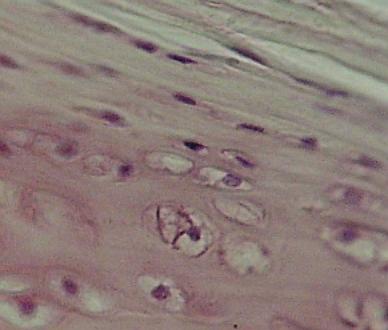
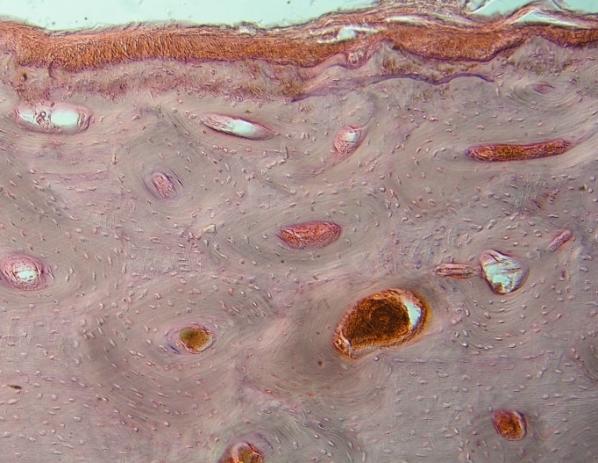
Hyalinní
chrupavka průdušnice člověka – shora ochrustavice (kolagenní vazivo), uprostřed
– fibroblasty se mění v chondroblasty, dole zralé chondrocyty
s velkým množstvím mezibuněčné hmoty – chondrinu. Kolagenní vlákna nejsou
zřetelná – stejný index lomu světla jako chondrin. VP. Srovnej též (URL
22)
Hyalinní
chrupavky larvy čolka s úpony svalových vláken kosterního svalstva. Po
obvodu chrupavky vznikají z fibroblastů chondroblasty (růst apozicí).
Chrupavka je mladá, ještě nevznikly izogenetické skupiny dělením chondrocytů
uvnitř tkáně. U vyvíjejících se chrupavek tvoří perichondrium zatím jen jemné
vrstvy (zleva i pod hrupavkou). V dolní části přechází perichondrium
v jemné epimyzium a časem se zde vytvoří šlacha.
c) Elastická (pružná) chrupavka - mnoho elastických vláken v základní hmotě, ta má jiný
index lomu, proto je na preparátech viditelná. Méně průsvitná, nažloutlá a
silně pružná (ušní boltce, ve stěnách velkých tepen, epiglotis).
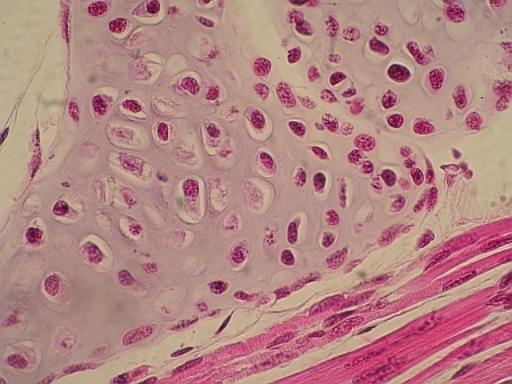
Elastická
chrupavka. Nahoře modře zbarvené vazivo perichondria přechází přes vrstvu
chondroblastů v hotovou tkáň tvořenou chondrocyty a jejich izogenními
skupinami.
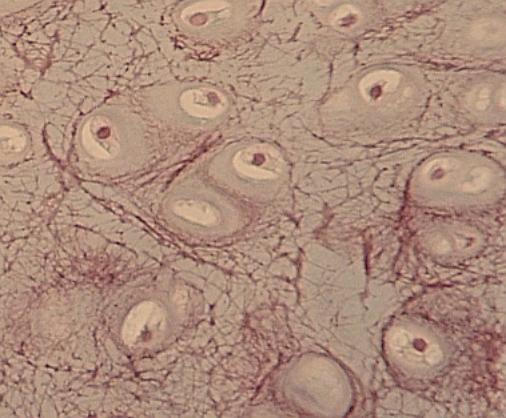
Detail
elastické chrupavky. Dvojice chondrocytů vzniklých dělením jsou obklopeny dobře
zřetelným teritoriem vláken, z nichž elastinová jsou dobře viditelná (mají
jiný index lomu než chondrin). Srovnej též (URL
22).
d) Vláknitá chrupavka kolagenní (vazivová) - méně chondrocytů
i interfibrilární hmoty, hojnost kolagenních vláken (meziobratlové ploténky,
čéška, kloubní plošky). Je průsvitná, bílá, velmi hutná.
Cartilago – chrupavka – vazivová - fibrocartilago
![]()
5.12. Kost
Počínaje
kostnatými rybami tvoří kostru obratlovců, tj. oporu a ochranu orgánů. Je
součástí pohybového aparátu. Původně chrupavčitá kostra je nahrazována kostní
tkání. Kromě vnitřního skeletu vytváří šupiny ryb, kožní desky krokodýlů a
želv, může vyztužovat některé měkké orgány (penis některých savců, jazyk
některých ptáků). Charakteristický je rozvoj mezibuněčné hmoty, která obsahuje
asi 25% organické složky, 55% anorganických látek a asi 20% vody. Organickou
složku tvoří GAG (glykosaminokany a vlákna kolagenu - ossein = kolagenní vlákna
+ glykoproteiny s kys. chondroitinsírovou B). Kolagenní vlákénka mají svou
úlohu při mineralizaci, na jejich povrch se ve formě plochých šupinek ukládají
soli Ca - fosforečnan a uhličitan vápenatý, (CaF2,
hydroxiapatit = Ca10(PO4)6(OH)2), hořčíku ve
formě fosforečnanu a NaCl. (Též Mg(PO4)2, SrSO4).
Kostní buňky. Z buněk rozlišujeme osteoblasty uložené na povrchu kosti - aktivně
syntetizují mezibuněčnou hmotu. Osteocyty = vyzrálé kostní buňky
definitivní kosti. Leží ve zvápenatělých komůrkách v základní hmotě a dotýkají
se svými cytoplazmatickými výběžky, které v mezibuněčné hmotě takto
vytvářejí droboučké kanálky. Na povrchu je kost kryta vazivovou okosticí
= periostem, z vnitřní strany endostem. Jako orgán má kost dvě
složky: kostní tkáň a kostní dřeň. Zvláštními buňkami dočasně se vyskytujícími
v kostní tkáni, jsou tzv. osteoklasty = velké, mnohojaderné buňky,
přítomné jsou při odbourávání chrupavky při její přeměně v kost nebo i při
odbourávání primární primitivní kosti při přeměně na kost definitivní.
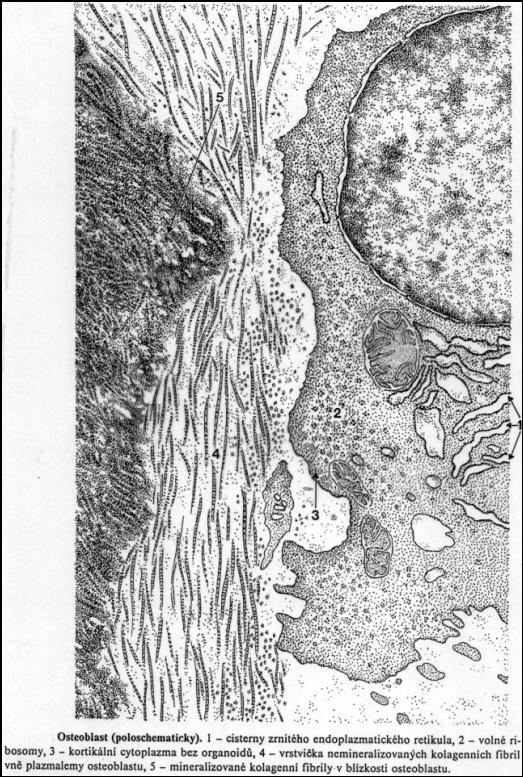
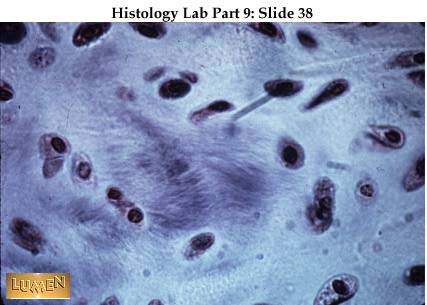
Osteoblast v EM
Osteoblast v EM. Převzato z Klika a
kol, (1986).
5.12.1. Primární (vláknitá) kost
je prvotní tkáň vzniklá v prvních fázích
osifikace. Osteocyty jsou zde
uloženy nepravidelně, mezibuněčná hmota obsahuje velké množství plsťovitě
propletených kolagenních vláken. V dospělosti bývá přítomna u nižších
obratlovců, obojživelníků a plazů. Jinak bývá nahrazována kostí definitivní –
lamelózní.
Ostenoidní tkáň - podobá se vláknité kosti. Setkáváme se s ní u některých kostnatých
ryb a u vyšších obratlovců je z ní tvořena zubovina, případně tzv. drsnatiny na
jiných kostech.
5.12.2.
Kost definitivní (lamelózní)
bývá uspořádána do lamel, které mohou vytvářet tzv.
kompaktu nebo kost houbovitou.
Kost kompaktní - v diafýzách dlouhých kostí a na povrchu kostí plochých. Stavba – z osteonů
(Haversovy kanálky a lamely - 8-16 lamel kolem Haversova kanálku, střídání
vrstev kostních buněk a mezibuněčné hmoty - nejrůznějšími směry). Je doplněna
systémem lamel plášťových vnitřních a vnějších. Lamely vmezeřené
– zbytek předchozích osteonů před poslední rekonstrukcí. Diafýzy jsou uvnitř
duté, vyplněné žlutou dření kostní( tukové pojivo). Haversovy a Volkmanovy
kanálky: jimi vnikají do kostí cévy a nervy.
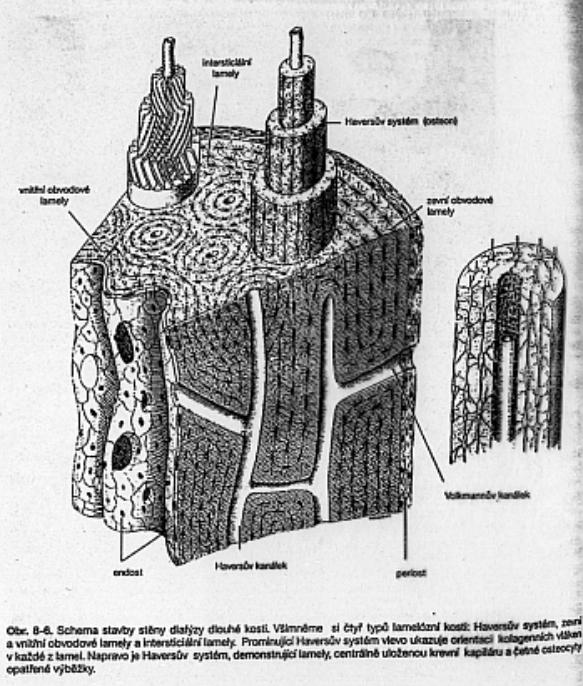
Kost lamelózní, kompaktní. Periosteum,
vnější plášťové lamely, Haversovy systémy, vmezeřené lamely, vnitřní plášťové
lamely endosteum (Janquiera a kol.)
Lamelózní kost kompaktní
s Haversovými systémy, vmezeřenými lamelami, a na povrchu krytá okosticí
(tuhé vláknité pojivo plsťovité), které proniká do povrchových vrstev kosti. VP
Kost
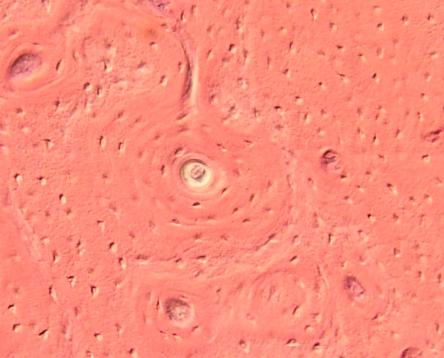
lamelózní, kompakta, čelist psa. Haversovy systémy s Haversovými lamelami,
vmezeřené lamely, Volkmannův kanálek (svislý proužek nahoře) VP
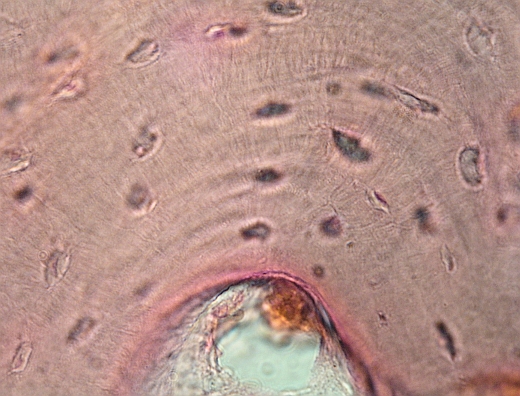
Haversův
systém při větším zvětšení, kanálek dole. Uvnitř
kanálku řídké vláknité pojivo, cévy. Koncentrické Haversovy lamely,
osteocyty tmavé, částečně jsou zřetelné jejich vzájemné plazmatické spojky
v kanálcích v kostní hmotě. VP

Schéma
téhož, co je nahoře.
Kost houbovitá - spongiózní je tvořena prostorovými trámci a ostny (spiculi)
lamelózní kosti, mezi nimiž je červená kostní dřeň. V některých silnějších
trámcích jsou osteony (výživa - kanálky v osteonech, trámečky jsou uspořádány
tak, aby nejlépe odolávaly tlaku), slabší trámce osteony nemají. Je
v epifyzách (hlavicích) dlouhých kostí, ve střední části plochých kostí a
tvoří kosti krátké.
Okostice - periost – pevný vazivový obal protkaný nervy a cévami, které
prostupují kost díky existenci Haversových a Volkmanových kanálků. Na vnitřní
straně diafýz je podobná vazivová vrstva - endost (zahuštěné pojivo kostní
dřeně), periost připoután ke kosti tzv. Sharpeyovými vlákny.
Kostní dřeň (morek) = síťovité pojivo retikulární. Obsahuje:
retikulární buňky, retikulární vlákna, tukové buňky a buňky, ze kterých se
diferencují erytrocyty, granulocyty a trombocyty.Tvorba krvinek = v
červené kostní dřeni,
Žlutá dřeň - tuk = degenerace ve stáří (šedá dřeň)
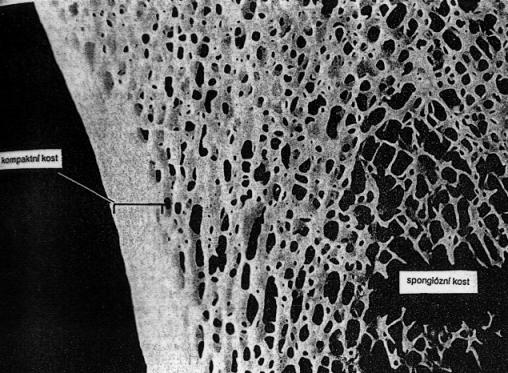
Kost lamelózní - kompaktní i houbovitá. Stavba dlouhé kosti v místě přechodu diafýzy v epifýzu. Vrstva kompakty vně,
houbovitá (spongiózní) kost uvnitř, směrem dolů dutina. V spongiózní kosti
červená kostní dřeň, v dutině dlouhé kosti tukové pojivo (žlutá kostní
dřeň).
![]()
5.12.3. Osifikace - u
kostí krycích (dermálních), které se diferencují přímo ze zárodečného
mezenchymu nebo z vaziva probíhá tzv. desmogenní osifikace (spodní
čelist, kosti lebeční, kostní desky v kůži želv a pásovců, krokodýlů). Vznik
tzv. kostí primárních - později je možná přestavba na kosti lamelózní.
Na povrchu osifikující tkáně se řadí mezenchymální buňky na způsob epitelů,
posunují se dovnitř a mění se na osteoblasty - ty vylučují mezibuněčnou hmotu a
mění se v osteocyty. Vznikne houbovitá kost, která později roste apozicí
(diploe).

Desmogenní
osifikace čelisti člověka. Na již hotové kostní trámečky (růžově fialové
s osteocyty) nasedají osteoblasty kubického tvaru vzniklé
z embryonálního mesenchymu, který se kromě v osteoblasty mění také
v retikulární pojivo, vyplňující světle zbarvené prostory. VP
Chondrogenní osifikace na místě hyalinní chrupavky je dvojího typu:
Perichondriální - začíná uprostřed diafýz pod ochrustavicí (perichondriem).
Buňky přítomné v pojivu ochrustavice se
diferencují v osteoblasty, které vytváří po obvodě diafýz prstenec kompaktní
kosti, tvořený nejprve jednou lamelou. Ochrustavice se mění v periost a
následná ossifikace probíhá přikládáním dalších lamel, každá je ve směru
diafýzy delší než předchozí. V dospělosti poslední povrchová lamela spojí
povrch diafýzy s povrchem epifýz.
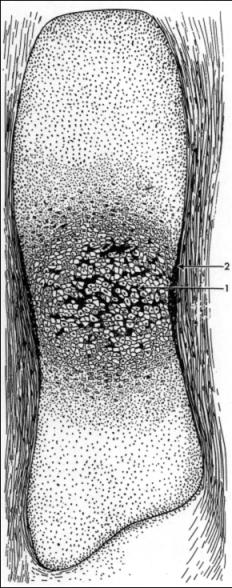

Vznik kosti – osifikace chondrogenní
na bázi chrupavky - enchondrální – zevnitř
,
perichondrální - zevně, z ochrustavice (Podle Klika – nahoře a Janquiera a kol.)
Enchondrální osifikace - přibližně v úrovni první perichondrální lamely se
v nitru diafýzy začne chrupavka kalcifikovat = chondrocyty se zvětšují a
v mezibuněčné hmotě se ukládají soli Ca2+. Do
zvápanatělé chrupavky vnikají z periostu pupence cév a mezenchymu. Z
mezenchymu vznikají prvně chondroklasty - velké mnohojaderné buňky,
které narušují chrupavku, fagocytují odumřelé chondrocyty a uvolňují další
místo pro cévy a mezenchym (směrové trámce). - Tak vzniká uprostřed diafýzy centrum
osteogenní tkáně s vlastním krevním zásobením a schopnosti diferenciace v houbovitou kostní tkáň. Od centra se proces šíří oběma sněry diafýzou a
později k němu dojde i v epifýzách. V době dospívání jsou chrupavky jen mezi
epifýzami a diafýzami (epifýzodiafyzální ploténka) a na povrchu epifýz, kde
tvoří kloub. Chrupavčitá zóna = růst kostí do délky. Rostoucí prvotní kost =
hrubě vláknitá spongiózní kost, kompaktní jen z povrchových lamel). V poslední
fázi, či bezprostředně dojde k postupné resorbci této tkáně a jejímu nahrazení tkání
jemně vláknitou – lamelózní. Osteoklasty rozrušují kost a vytvářejí prostory
pro její přestavbu - široké dutiny =vstup osteogenní tkáně = Haversův systém
lamel. Vmezeřené lamely = zbytek po dřívějších Haversových lamelách, které při
růstu kostí byly také částečně odbourány.
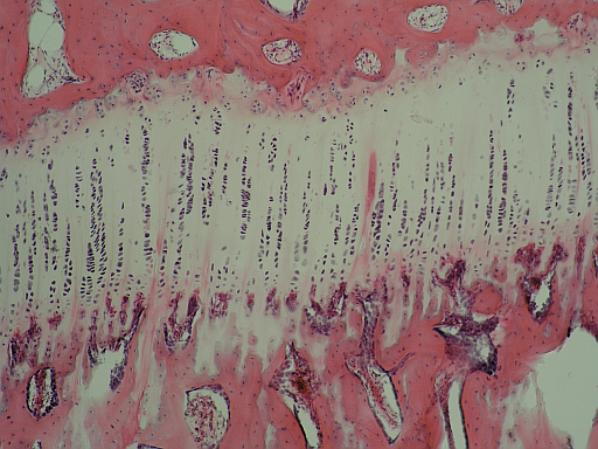
Enchondrální ossifikace. Nahoře růžově
zbarvené trámce houbovité kosti v epifýze, uprostřed světlý pás
epifýzo-diafýzální chrupavčité ploténky (shora: zóna normální chrupavky, zóna
růstu s vertikálními řetízky izogenetických skupin chondrocytů, níže zóna
kalcifikace chrupavky, pod ní zóna eroze chrupavky makrofágy (chondro- nebo též
osteoklasty, růžové „kuličky“ uvnitř světlé masy zbytků chrupavky). Následuje
zóna ossifikace, kde z retikulárních buněk se vyvíjejí vrstvičky osteoblastů,
které se postupně mění v osteoklasty a dávají vznik primární (hrubě
vláknité) houbovité kosti, jejíž trámečky postupným přikládáním dalších vrstev
osteoblastů a obalování osseinem postupně tloustnou (zcela dole). Uvnitř
houbovité kosti je krvetvorná červená kostní dřeň (retikulární pojivo,
z jehož kmenových buněk vznikají krevní buňky, případně osteoblasty
dalších generací).
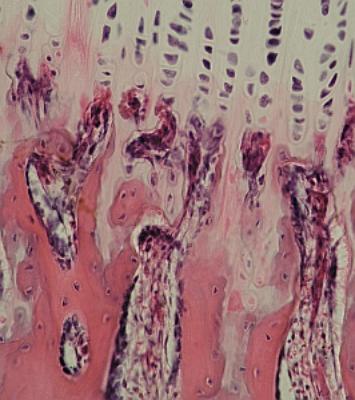
Ossifikace
enchondrální při větším zvětšení. Linie makrofágů (chondroklastů) přibližně
v horní třetině. Jsou nasměrovány proti řadám chondrocytů, nikoli jen tak
náhodně do chrupavky nahoře.
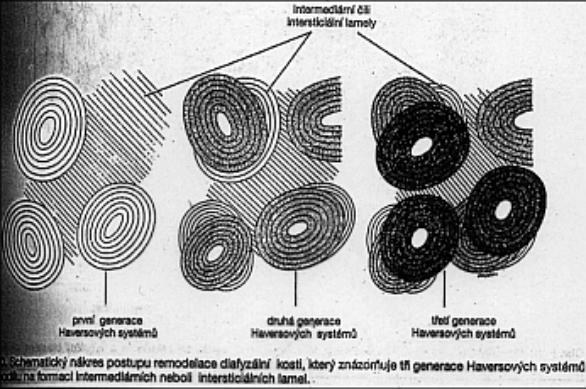
Schéma
lamelózní kosti po několikeré přestavbě (Janquiera a kol.)
Přestavba kostí se děje pomocí makrofágů – osteoklastů, které původní tkáň rozpustí a vytvoří široký kanál, a osteoblastů, které se jako retikulární pojivo do kanálu nastěhují a postupně vytvářejí koncentrické lamely od obvodu směrem do středu. Po vytvoření poslední lamely zbude v centru Haversův kanálek.
Při růstu do délky se odbourává (osteoklasty) spongiózní
tkáň v nitru diafýz, zde však se nahrazuje jen žlutou kostní dření.
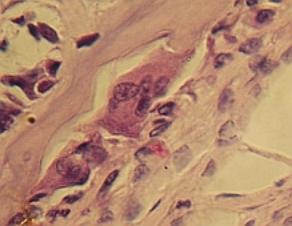
Dva
osteoklasty (přibližně uprostřed nad sebou) odbourávají primární houbovitou
kost (šikmo vlevo nad nimi) uvnitř budoucí diafýzy ulny člověka. Směrem vpravo
vrstva nediferencovaných buněk a zcela v pravo dole fibrocyty periostu
U ptáků se
do dutých kostí vchlipují plícní vaky = pneumatizované kosti.
Hojení zlomenin - z periostu, přes chrupavčitý tzv. svalek a jeho náhradu kostí
s následnou finální rekonstrukcí. Úplná regenerace odňatých kostí je možná
jen u nižších obratlovců. Kosti = původ z mezenchymu.
![]()
5.12.4. Spoje
kostrové
1) Synartrosis - nepohyblivé spoje vazivem či
vmezeřenou chrupavkou či kostní tkání bež štěrbiny:
a) synostosis
- spojení pomocí kosti - ossa illium + pubis + ischii = os coxae (kost
pánevní), os sacrum (křížová) = z pěti obratlů,
b)
synchondrosis - spojení pomocí chrupavky symfysis ossis pubis (srůst
kostí stydkých), nebo žebra a kost
hrudní (přes hyalinní chrupavku).
c)
syndesmosis - přes
vazivo (sutury lebeční, závěsný aparát zubu).
2) diartrosis - pohyblivé spoje, klouby s kloubní
štěrbinou, kloubní pouzdro z hustého kolagenního vaziva, synoviální mok.
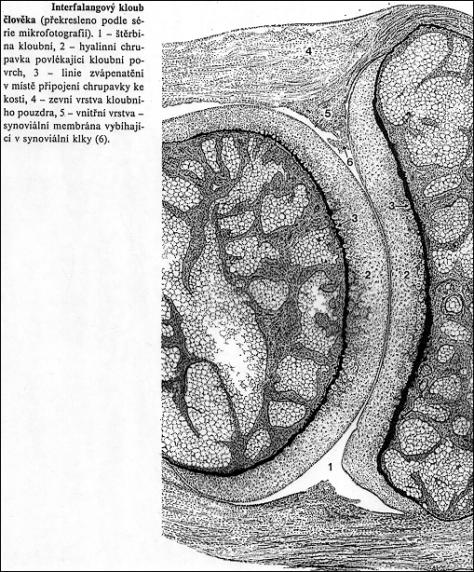
Kloubní spoj – diarthrosis - pohyblivý
Růst a diferenciace kostí – vliv endokrinní sekrece:
1) Hypofýza
2) Štítná žláza + příštitná tělíska
3) Placenta
4) Pohlavní žlázy – varlata i vaječníky
![]()
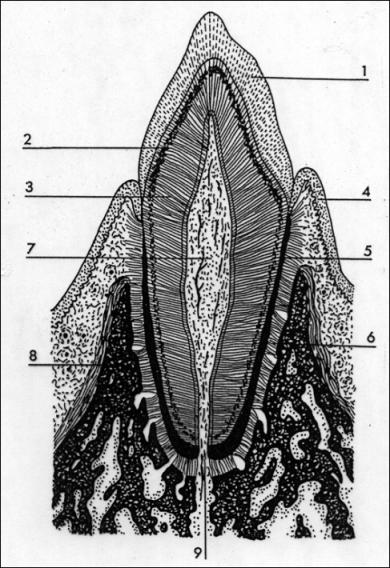
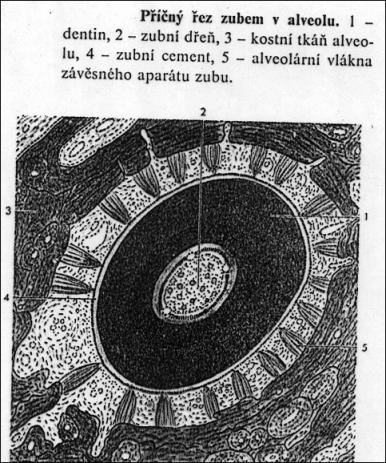
5.13. Zubní tkáně
Mají původ v mezenchymu a ektoblastu. Z krycího epitelu
vyvíjející se čelisti se vychlipuje souvislá lišta se dvěma řadami (v případě
dvojího chrupu) pohárkovitých útvarů tzv. sklovinných orgánů. Do dutin
těchto pohárků vniká mezenchymální tkáň. Obě složky vytváří základ zubu.
Horní sklovinné orgány vytváří mléčný zub, spodní zub trvalý. Sklovinný orgán
vylučuje sklovinu - substantia adamantina. Na povrchu mezenchymové
papily vycestují odontoblasty a seřadí se na způsob epitelu. Odontoblasty
vylučují dentin - zubovinu. Z odontoblastů vybíhají do dentinové hmoty
tzv. Tomesova vlákna – inervované cytoplazmatické výběžky odontoblastů.
Mezenchymová papila vyplňuje dutinu zubu (cavum dentis) a později se
diferencuje v zubní dřeň - pulpa dentis. Ta je tvořena řídkým vláknitým
až rosolovitým pojivem, nervy a cévami. V zubním lůžku (alveolus) čelisti je
zub upevněn tmelem (cementem) - substantia ossea, což je vláknitá kostní tkáň
na kořenové části zubu. Mezi cementem a kostí lůžka je vazivová blána (ozubice)
- periodontium s podobnou funkcí jako okostice. Obdobou zubní tkání jsou
plakoidní šupiny a hákovité zuby na čelistech žralokovitých.
Email - 98% anorg. látek (krystaly hydroxiapatitu)
+2% organických látek (převážně kolagenní vlákna)Prismata - průměr 5-10 um od
povrchu skloviny k dentinu. Průběh - radiálně spirálně, radiálně (zatížení
při žvýkání). Povrch emailu - aprismatická zóna (cutiata dentis -org. vrstvička částečně chybí či
zvápenatělé povlaky).
Dentin - 72% org. látek - fosforečnan vápenatý
(hydroxiapatit)
28% org. látek. Průběh: Dentinové kanálky (tubuli
dentis) radiální (dole) a esovitý (nahoře).


Vše podle Kliky a kol., (1986)
Viz též URL 29
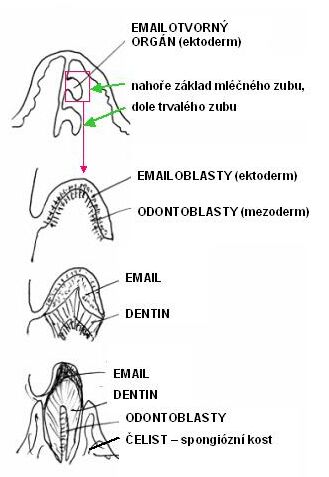
Schéma embryonálního
vzniku zubu (v příčném řezu tvořící se dásně)
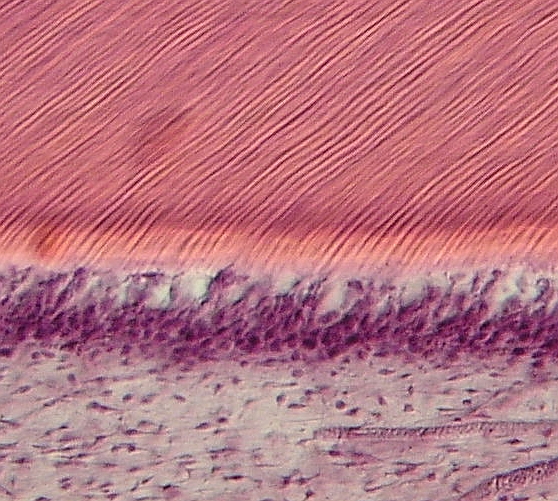
Dentin
(nahoře) a pulpa dentis (dole) v zubu psa. Zřetelná je vláknitá struktura
zuboviny, tvořená jemnými kanálky kolem cytoplasmatických výběžků odontoblastů,
které sahají až k hranici dentinu. V kanálcích jsou též nervové
výběžky. Odontoblasty samy tvoří vrstvu buněk těsně pod dentinem. Samotná pulpa
je typem rosolovitého pojiva. VP
![]()
5.14. Pojiva trofická
(tělní tekutiny)
1) Rozvádění živin, výměna plynů, obrana organismu,
rozvod hormonů. Podobají se co do složení a funkce řídkým vláknitým pojivům.
2) Kolují v mízních a krevních cévách nebo se
rozlévají v tělních dutinách a buněčných prostorách.
3) Mezenchymový původ (vývoj jiné přes pojiva)
4) Různé typy volných buněk
5) Tekutá mezibuněčná hmota
6) Krev, krvomíza, míza, tkáňový mok
5.14.1. Míza - krevní
plazma (podobná krevní plazmě, více tukových látek, mléčně zkalená). Málo O2, mnoho CO2.
- lymfocyty (asi 90%) buněk mízy -
malé, střední a velké
- monocyty + malé množství erytrocytů a
eosinofilních granulocytů.
Funkce -
vyživovací, obranná (ale u některých bezobratlých i přenos O2 - krevní barviva rozpuštěná). - Ductus thoracicus - u
savců, u člověka průměr 4-6mm.
5.14.2.
Krev - tělní tekutina červené barvy
kolující v systému uzavřených cév. Poháněna srdcem nebo silnější
z cév. U člověka množství 5,5-6 litrů
Funkce -
rozvádění živin, výměna plynů mezi tkáněmi a dýchacími orgány, odstraňování odpadních
látek do vylučovacích orgánů, rozvádí hormony, obrana těla, udržování tělesné
teploty.
Velké rozdíly mezi krví obratlovců a bezobratlých.
Krevní plazma - hustá, nažloutlá, 9O% H2O +
minerálních a organ. látek: Na+, Mg2+, K+, Ca2+, Cl-, I-, SO42-, PO43- O2, CO2 N, glukóza O,1%, bílkoviny 7%, zbytek kapénky tuku
(chylomikrony), mastné kyseliny a celá řada org. látek.
Poměr buněk a plazmy = hematokrit (odstředění v
kapiláře, muž 39-49%, žena 35-43%)
Obsah minerálních látek: 0,85% teplokrevní,
0,65% studenokrevní,
<3% mořští bezobratlí
Bílkoviny
- sérumalbuminy - trombogen, sérum globuliny - enzymy, alfa-, beta-, gama-
fibrinogen, achroglobuliny (chromoproteiny) funkce krevního barviva u některých
bezobratlých.
Krvinky - obratlovci: erytrocyty (obs. hemoglobin)
leukocyty, trombocyty:
Erytrocyty u savců bezjaderné, ploché, bikonkávní, (s výjimkou velbloudovitých,
kde oválné), jinak ploše oválné, kromě savců s jádrem (člověk průměr 7,5 um).
Počet :
skokan 250.000, člověk 4-5 miliónů, kapr 1.9 mil.
koza 13-17 miliónů, kůň 6-9 mil. v 1 mm3.
Polyglobulie = více erytrocytů než norm. hodnota
Anemie (oligocytemie) - méně než norm. hodnota
erytrocytů
Hlavní funkce erytrocytů = přenos kyslíku
Hemoglobin
- až 95% sušiny erytrocytů
- hemochromogen + Fe + globin (albumin)
- hemoglobin + O2 -
oxihemoglobin
Životnost u člověka 14 - 120 dnů.
V hypotonickém roztoku: hemolýza.
Leukocyty -
počet v 1mm3: člověk 6-10 tisíc, kapr 15 tisíc
koza 8-16 tisíc, kůň 7-12 tisíc
a) Pravé – granulocyty - velké
jádro různého tvaru, mnoho ergastoplazmy (zrnitost). Vznik v
kostní dřeni
- neutrofilní - mnoho drobných granulí v
cytoplazmě, špatně barvitelné kyselými a zásaditými barvivy. Mladá forma
neutrofilu (segmentu) tyčka (jádro nesegmentované, tyčinkovitého tvaru),
zaškrcované (zralé) = segment. Fagocytují drobné částice - mikrofágy. 60-70%
všech leukocytů. Infekce = zvýší se tyčky.
- acidofilní (eosinofilní) - velká granula
(kyselá fosfatáza, katepsin, ribonukleáza) dobře barvitelná kys. barvivy
(Giemsa - oranžově až sytě červeně).
Jádro
(tvar činky, sluchátka, brýlí). 2-4% všech leukocytů. Pomalá fagocytóza,
amoeboidní pohyb, obsahují protofibrinolyzin (udržuje tekutost krve).
- bazofilní granulocyt (bazofil) velká, hrubá
granula, Giemsou barvitelné modře, kryjí jádro. Jádro téměř kulovité,
nepravidelné. U člověka 1% všech leukocytů. Cytoplazma obsahuje heparin
(zábrana srážení krve při zánětech).
b) Agranulocyty -
vznik v lymfatickýxh tkáních, prekurzory v kostní dřeni. Jádro
oválné, kulovité, ledvinovité. Cytoplazma bez granulí.
- lymfocyty - v krvi jen malé, v kostní dřeni
a míze malé, střední a velké, (někdy v krvi i větší a střední). Zejména ve
tkáních se záněty (nejvíce na konci infekčního období), jinak množství 20-30%
(u člověka) leukocytů. Asi (i) produkce protilátek. TL = z brzlíku (thymus), BL
´ bursa Fabricii (ptáci), jinak apendix, tonsily, kostní dřeň. Životnost 20-25
dnů, ale i dlouhožijící, paměťové, buňky.
- monocyty - největší krevní
buňky (16um). Jádro - velké,
ledvinovité, excentricky umístěné. Makrofágové.
6-8% leukocytů (u člověka). Hojnější při infekcích a zhoubných leukemiích.
Vznik v kostní dření, zdroj jiných forem makrofágů.
Trombocyty (krevní destičky) - okrouhlé destičky cytoplazmy bez jádra (4um), jen u
ptáků samostatné buňky podobné erytrocytům. U savců vznikají v kostní dřeni z tzv. megakaryocytů.
Po poranění uvolnění tromboplastického faktoru = srážení krve.
Tromboplastické faktory trombocytů a poraněných buněk + jonty Ca2+
změní protrombin na trombin, ten pak katalyzuje změnu - fibrinogen – fibrin – vláknitá síť pro zachycení krevních buněk
a ucpání rány.
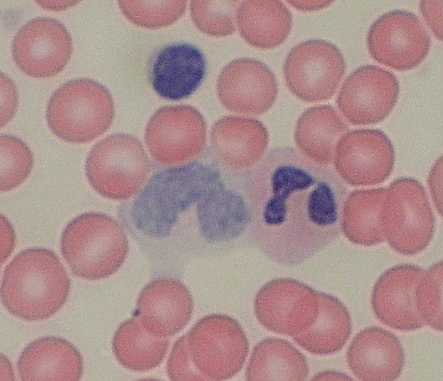
Krev
člověka – erytrocyty, nahoře lymfocyt, dole uprostřed monocyt (makrofág) a
neutrofilní granulocyt (mikrofág). VP
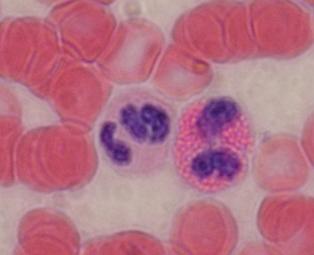
Neutrofilní
(vlevo) a eozinofilní granulocyt
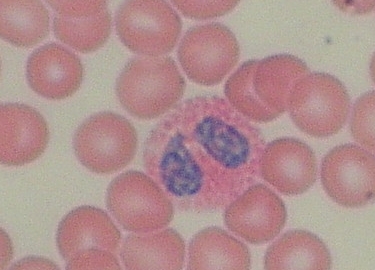
Eozinofilní
(acidofilní) grynulocyt s typickým tvarem jádra.
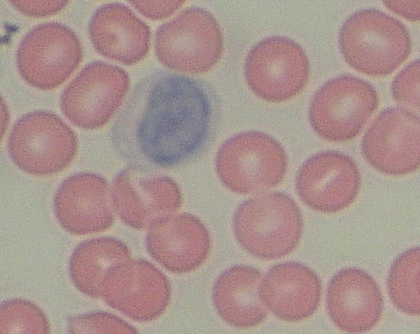
Monocyt
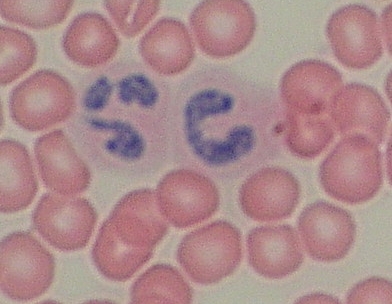
Neutrofilní
granulocyty – starší se segmentovaným (ale spojené segmenty!!!) jádrem a
mladší, tzv. „tyčka“ s jádrem dosud kompaktním. VP
Pozor, v ukázkách
schází bazofilní granulocyt – cytoplazmu má vyplněnou granulemi v uvedeném
typu barvení by byl celý modrý, jádro maskováno cytoplazmatickými zrny.
Monocyty
v krvi myši - fagocytóza kuliček z plastu. Foto P. Hyršl
Krvetvorná tkáň v červené kostní dření při
enchondrální ossifikaci lidské ulny. Uprostřed je retikulární krvetvorné pojivo
s buňkami v různém stadiu diferenciace. V horní třetině
uprostřed je velký megakaryocyt (budoucí krevní destičky). Oblasti se zralými erytrocyty
jsou cévy, nejzřetelnější je vpravo dole. VP
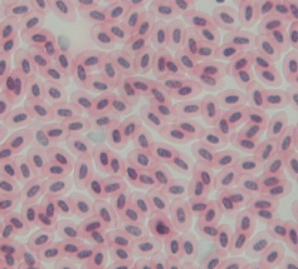
Krev ptáků. Erytrocyty mají jádro. Podobně vypadají
tzv. koagulocyty, odpovědné za srážení krve.
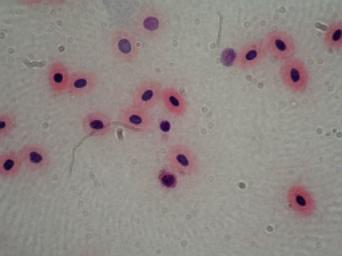
Krevní buňky kapra (Cyprinus) mají jádro. VP
Zde (URL
31) najdete krátké video jak se dělají krevní roztěry (Smear Examination),
a také krevní buňky různých živočichů (Blood Cells by Species)
![]()
5.14.3. Teorie
vzniku krevních buněk
Bezobratlí
– krevní buňky vznikají z mezenchymu a pak se množí přímo v hemolymfě.
Obratlovci
- z počátku podobně jako u bezobratlých, ale později se krvetvorba omezuje
(soustředí) do tzv. krvetvorných center.
Krvetvorná centra u dospělých:
Kruhoústí - pojivo střeva.
Ryby a obojživelníci – slezina.
Ptáci a savci - v embryogenezi žloutkový váček, pak játra, jinak ve
slezině, mízních uzlinách a kostní dřeni - hematopoetické orgány. Rozeznáváme:
orgány myeloidní - (erytrocyty agranulocyty, monocyty) kostní dřeň
a
orgány lymfoidní - (lymfocyty) - prekursory
kostní dřeň, pak množení v lymfatických orgánech (slezina, brzlík, bursa
Fabricii, lymf. uzliny).
Schéma vývoje krevních buněk člověka.
Převzato z http://www.lrf.org.uk/en/1/infdispatbmt.html
5.14.4.
Tkáňový mok – nachází se ve všech
tkáních mezi buňkami. Vytváří jejich životní prostředí a zajištuje mezi nimi
výměnu látek prostředí všech tkání. Produkce - buňky krevních vlásečnic a buňky
tkáňové. Složení proměnlivé. Méně bílkovin než krevní plazma.
5.14.5. Mozkomíšní mok – podobný
moku tkáňovému, minimálně buněk (lymfocyty), produkován ependymem dutin
v centrální nervové soustavě.
5.14.6.
Krvomíza – hemolymfa u
hmyzu a jiných bezobratlých . Ze srdce přímo do tělních dutin a mezibuněčných
prostor ve tkáních. Koluje v neuzavřených cévních systémech.
Plní funkci lymfy i krve. Volné buňky schopné fagocytózy. U hmyzu bez krevních
barviv (tracheální soustava), jinak obsahuje barviva pro vazbu kyslíku.
5.14.7. Hemocyty hmyzu
Klasifikace je velmi variabilní a není ujednocena
systematika. Podle Jonese existují:
1) Prohaemocyty - malé, kulaté, poměrně velký nucleus
a silně bazofilní cytoplazma, často se dělí a jsou předchůdci jiných typů.
2) Plasmocyty - často nejpočetnější typ, bazofilní
cyto - plazma, variabilní formy, fagocytóza (jádro má být asi o 15%
větší), nebo enkapsulace.
3) Granulární haemocyty - fagocyty s acidofilními
granulemi
v cytoplazmě.
4)Cystocyty
(koagulocyty) - ve fázově kontrastním zorném poli mají hyalinní cytoplazmu,
ostře difinované malé jádro, asi modifikovaný gonolární? haemocyt.
U některých druhů hmyzu:
5) Oenocyty - Coleoptera, Lepidoptera, některá
Diptera a Heteroptera. Velké bazofilní buňky s granulemi nebo s krystaly v
cytoplazmě.
6) Spherule cells (buňky se sférickými – kulatými –
útvary v cytoplazmě) - Diptera, Lepidoptera.
Velké oválné buňky s acidofilními inkluzemi.
7) Adipohaemocyty - Diptera, Lepidoptera + někteří
jiní ze studovaných. Kapičky tukové povahy v cytoplazmě.
Larvy Musca - žádné haemocyty v lymfě, larvy
Periplaneta - mnoho haemocytů v lymfě.
Funkce: sekreční činnost, stimulace prothorakálních
žláz, tvorba tukového tělíska, výživa tkání, likvidace poranění (melanizace)
nebo parazitů (enkapsulace).
Hemolymfa hmyzu
Hemocyty bource
morušového
Prohemocyt
Sferulocyt
Oenocytoid
Granulocyty (d,e,f)
Plasmatocyty (g,h,i)
Adipohemocyty
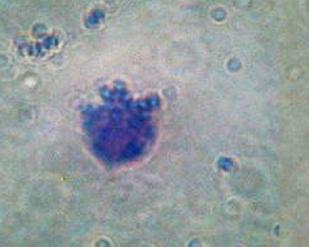
Hemocyt
hmyzu fagocytující kuličky z plastu. (P. Hyršl)
![]()
5.14.8. Krevní barviva
Hemoglobin
(krevní barvivo obratlovců, ale i některých bezobratlých - kroužkovci)
- bílkovina globin 96%
- nebílkovinný pigment hem 4% (protoporfyrin) s Fe2+
Hb + O2 = HbO2 (oxihemoglobin)
HbO2 = O2Hb = redukovaný hemoglobin
Plná saturace Hb - když je všechen vázán na O2
(200ml O2 v 1 l krve)
CO2 + Hb -
karbominohemoglobin (slabá vazba)
O3 + Hb - methemoglobin
silné oxid. činidlo - stálý (Fe3+)
Hb + CO - karboxihemoglobin (stálý) již O,1% CO je
nebezpečné (210krát více váže se než O2.
Chlorokruoriny - mořští červi - Fe
Hemerytriny – Sipunculidae - Fe
Erytrokruoriny – pakomáři – Fe
Echinochrom – ježovky - Fe
Hemocyanin
- (bílkovina obsahující Cu) - rak, škeble, hlemýžď, hlavonožci (jen asi 70ml v
1 l krve).
Achroglobin – V (vanad –
pláštěnci, bez barvy).
|
Krevní barviva: složka
bílkovinná a nebílkovinná Hemoglobiny Fe (červené: obratlovci, někteří kroužkovci, korýši, pakomáři
Chironomus aj. bezobr.) (OxiHb, karbaminoHb,
metHb, karboxiHb) (A Hb
–dospělců, F Hb – plod) Hemerytrin Fe (fialový: sumýšovci,
kroužkovci) Erytrokruorin Fe (červený: někteří pakomáři,
kroužkovci) Echinochrom Fe (žlutočervený: ježovky) Chlorokruorin Fe (zelený: někteří mořští kroužkovci) Hemocyanin Cu (modrý: škeble, hlemýžď, hlavonožci) (rak?) Achroglobin
Va (bezbarvý: pláštěnci) Rozdíl mezi krví obratlovců a bezobratlých: Bezobratlí mají
dýchací barviva rozpuštěna v krevní plazmě, obratlovci je mají v
specializovaných krevních buňkách. |
Kromě výše
uvedených tekutin vyskytují se v živočišné říši ještě
tekutina
coelomová, vyplňující coelomové
váčky kroužkovců a
tekutina perienterická, která vypňuje pseudocoelom hlístic a jiných
organismů s podobnou organizací.
![]()
6. TKÁNĚ SVALOVÉ - specializace buněk na pohybovou funkci
V cytoplazmě - myofibrily.
Popudy k činnosti - odstředivé nervové dráhy, nebo tkáňové hormony.
Typy:
svalové epitely
hladká
svalovina
žíhaná
svalovina
srdeční
svalovina
6.1. Myoepitely - přeměnou buněk krycích nebo
výstelkových epitelů - podkožní svalovina láčkovců, okružní svalovina a
podélná střevní svalovina u ploštěnců. Modifikace svalovina i epitel. -
velké svalové buňky Nematod. Obratlovci - oční duhovka, (zužování zornice) a
některé žlázy – vyprazdňování váčků. Původ - ektoblast či entoblast.
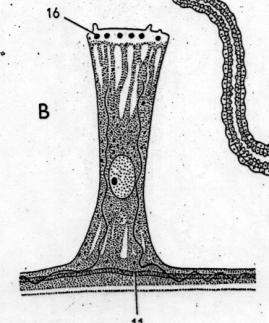
Buňka myoepitelu v ektodermu nezmara. 16 - vnější strana buňky, 11 (dole) výběžek se
stažitelným vláknem (podle Dogiela)
6.2.
Hladká svalovina - svalová buňka myocyt. Vřetenité, vláknité, někdy roztřepené,
15-200um.
a) Sarkolema - na povrchu
b) Jádro - oválné, protažené,
při stahu se může spirálně stáčet
c) Sarkoplazma
- s myofibrilami, sarkozómy (mitochondrie) a endoplazmatické retikulum drsné i
hladké, myoglobin
d)
Diplocentrum (obvykle 2 dělící tělíska)
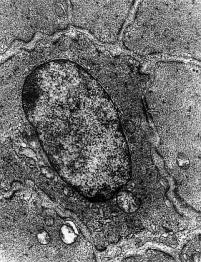
EM – řez
sarkocytem (Klika a kol, 1986). Uprostřed jádro, kolem buňky slabé endomysium
(ŘVP)

Schéma ukotvení
aktinových mikrofilament v sarkocytu. Densní tělíska jsou cytoskeletární
struktury ukotvené buď v cytoplazmě nebo pod CPM. Při stahu myocytu se
jádro může spirálovitě stočit.
Myofibrily
opticky 1 lomné - nežíhané. Sarkocyt se může stáhnou až o 1/2 . Bílkoviny ve
fibrile: aktin, myozin, tropomyozin, paramyozin. Funkce dosud ne zcela
objasněna. (Teorie: mikrofilamenta jsou zakotvena do tzv. denzních tělísek
v cytoplasmě nebo při plasmatické membráně tvořených intermediálními
filamenty. Za přítomnosti proteinu kalmodulinu a iontů vápníku dojde
k vzájemné reakci aktinu a myosinu a buňka se stahuje.) Vznik -
embryonální mezenchym – přes myoblasty. Sarkocyty jsou schopné se dělit a mají
základní funkční organely živočišné buňky.V dospělosti jde o tkáň smíšenou
s retikulárním pojivem (růst a obměna) až řídkým vláknitým pojivem – ochrana,
vascularizace. U obratlovců - vegetativní svalovina, vůlí
neovlivnitelná, jen pod vlivem vegetativního nervstva, Podráždění – mediátory
z nervových zakončení nebo ze sousedních svalových buněk. Jeden neuron
neinervuje všechny buňky ve skupině. Bezobratlí (mají jen hladkou
svalovinu kromě členovců a ploutvenek) -Je ovládána nervstvem
(svalovina mozaiková – jednotlivé buňky jsou inervovány - kroužkovci, měkkýši).
Paramyosinová hladká svalovina (mlžů) - stužkovité útvary paramyosinu kolem
nichž jsou uspořádána vlákénka aktinu a myosinu - schopnost dlouhé kontrakce za
malé spotřeby energie – svěrače lastur mlžů. Helikární svalovina např.
kroužkovci i jiní bezobratlí - prudká reakce. Obecně hladká svalovina –
dlouhodobá pomalejší činnost.
Hladký sval - stupňovitá
odpověď na podráždění (difuze mediátoru
a vedení podráždění od buňky k buňce.)
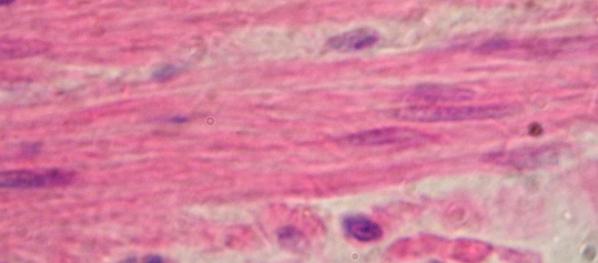
Hladká svalovina tenkého střeva člověka, podélně.
Vřetenovité myocyty mají podlouhlá jádra a jsou poměrně těsně u sebe, což bývá
pravidlem u obratlovců. Každý myocyt má samostatné endomysium, někdy však velmi
jemné. Svazky myocytů mohou být odděleny silnějšími vrstvičkami endomysia –
retikulárního až řídkého vláknitého pojiva, jako je to v tomto případě.
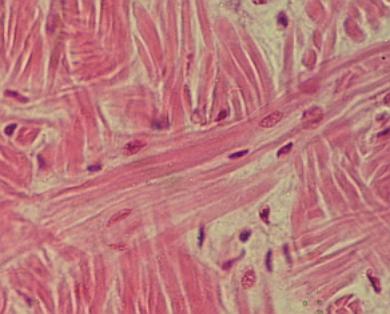
V různých
směrech uspořádané buňky hladké svaloviny nohy škeble (Anodonta sp.) Tmavá,
kompaktní jádra patří buňkám pojiva prostupujícího svalovinu
6.3.
Žíhaná svalovina
- fylogeneticky
mladší typ. Vzácně však i u primitivních mnohobuněčných nebo u členovců a
ploutvenek jako jediný typ svaloviny.
Obratlovci - kosterní svalstvo, lokomoční svalstvo, řízena CNS.
Základní jednotka - svalové
vlákno. Svalové vlákno = mnohojaderná buňka až několik cm dlouhý útvar krytý sarkolemou
- silná a pružná. V sarkoplazmě až několik stovek jader a množství
žíhaných vlákének - myofibril,
bohaté sarkoplazmatické retikulum ( hladké bez ribozómů - obaluje myofibrily,
slouží jako zdroj iontů Ca, možná se podílí na vedení vzruchu) a příčné tubuly,
které se sem zanořují jako vchlípeniny
cytoplazmatické membrány a vedou vzruchy z povrchu dovnitř svalového
vlákna. Velmi hojně jsou zastoupeny mitochondrie. Kyslík z okolních
kapilár přejímá v cytoplasmě hojně obsažený myoglobin (větší afinita ke
kyslíku než hemoglobin).
Jádro
svalového vlákna. Nahoře 3 – 4 žíhané myofibrily. Pod jádrem cytoplazmatická
membrána a kolagenní vlákna ŘVP) (Klika a kol., 1986)
Ultrastruktura
myofibrily –
telofragmy (příčné přepážky
z intermediálních filament umožňují zakotvení jednolomných vláken F
–aktinu, na která jsou vázány molekuly tropomyosinu a troponinu. Mezi aktinová
vlákenka částečně zasahují vlákna myozinu II. Místo mezi sousedními
telofragmami se nazývá sarkomera.
Svalové vlákno - stah:
Myosin - velké
množství molekul tvoří jedno vlákénko (viz cytoskelet v cytologii). Každá
z molekul má vláknitou část a hlavici. Hlavice je sídlo ATPázové
aktivity.
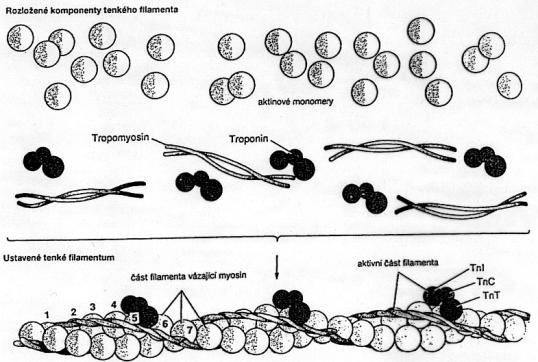
Aktinové
vlákénko (nahoře) - z mnoha kulovitých molekul G-aktinu, z nichž
vznikají dvě vlákna F-aktinu, která jsou spirálně stočena. K nim se váže
vláknitý tropomyosin a v určitých úsecích na okrajích tropomyozinu
nasedá regulační bílkovina troponin. Troponin brání vazbě hlavic myosinu
s aktinem. Zvýšením koncentrace Ca2+ iontů ,
které se vylijí z hladkého endoplazmatického retikula po podráždění
nervovým impulzem, změní troponin svoji prostorovou konfiguraci a odhalí na
aktinovém vlákně vazebná místa, které umožní vazbu myosinových hlavic. Ty se
váží na aktin (za spotřeby ATP) a změnou prostorové konfigurace (ohnutím
hlavice, opět za spotřeby ATP) posunují aktínová vlákénka směrem do středu
sarkomery. Další ATP molekuly umožňují disociaci hlavice myosinu od aktinu a
nové připojení v následujícím vazebném místě blíže tzv. Z linii –
středu zakotvení myozinových vláken. Tak je aktin postupně vsunován mezi
vlákénka myosinu. Činnost je současná a naráz. Myofibrila se stahuje. Klesne-li
Ca2+ koncentrace vyčerpáním vápníku do EPR, přestane myosin
reagovat s aktinem a působením ATP se vazby uvolní. Svalové vlákno se uvolňuje
(pružností cytoplazmatické membrány i předchozí spotřebou ATP). V nepřítomnosti
ATP relaxace nenastává (posmrtná ztuhlost - rozpad ATP ve svalech).
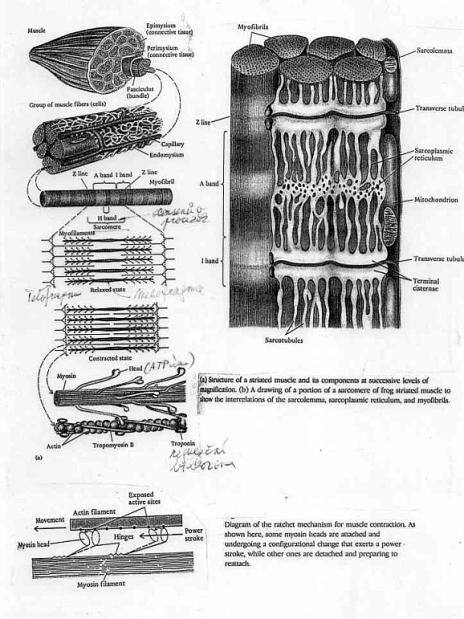
Schéma
žíhaného svalu (Barnes a kol.) od ultrastruktury myofibril (vlevo dole a uprostřed) přes svalové vlákno jako
jednotku žíhané svaloviny (uprostřed) až po sval (vlevo nahoře).
Svalové vlákno je obklopeno endomysiem z ŘV pojiva, snopce vláken jsou
obaleny perimysiem a sval kryje tzv. svalová povázka – epimysium,
z hustého vláknitého pojiva plsťovitého, které přechází ve šlachu, a ta
pak v okostici na povrchu kosti. Vpravo nahoře jsou myofibrily svalového vlákna
obklopené hladkým endoplasmatickým retikulem (cisterny pro Ca2+
ionty) a příčnými (tranzverzálními tubuli, které se vchlipují od povrchu CPM a
vedou depolarizaci CPM dovnitř svalového vlákna – jednorázové podráždění celé
struktury.
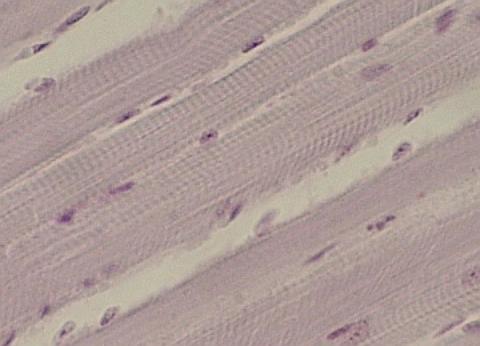
Žíhaná
svalová vlákna kamzíka (Rupicapra) s podlouhlými jádry pod povrchem – cytoplazmatickou
membránou. Každé je obaleno jemnou vrstvičkou endomysia – řídkého vláknitého pojiva. VP
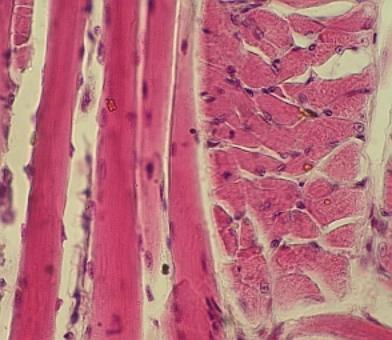
Žíhaná
svalovina jazyka kočky (Felis), řez
vlevo podélně, vpravo příčně (myofibrily ve vláknech jsou difúzně rozptýlené).
Kolem jednotlivých vláken se nachází jemné endomysium.
Žíhané
myofibrily jsou uspořádány ve vlákně buď rovnoměrně nebo tvoří tzv. Cohnheimova
políčka (sarkostyli). U žralokovitých ryb, u členovců a v srdeční svalovině
všech obratlovců jsou tzv. myotenie, páskovité myofibrily.
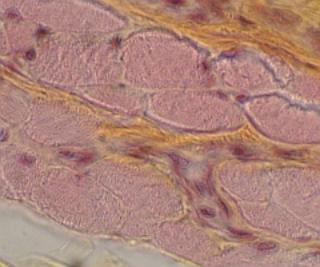
Příčně
rozříznutí svalová vlákna víčka člověka s difúzně rozptýlenými
myofibrilami. Vazivo endomysia a perimysia je zbarveno žlutě. VP
Struktura
svalu – svalové vlákno, obal z pojiva – endomysium (ochrana inervace, vascularisace),
více vláken obaleno perimysium a povrch svalu obaluje husté vláknité pojivo –
epimysium – fascie svalová, která se mění ve šlachu a upíná svaly na kostru –
do perichondria, periostu nebo na jiné struktury.
Svalovina
-
bledá (fázická): mnoho myofibril
méně myoglobinu a sarkozómů (mitochondrií) krátkodobě velká výkonnost.
-
červená (tonická): méně
myofibril, více myoglobinu a sarkozómů - dlouhodobě méně výkonná.
Střídají se v různých okrscích ale i převládají u
určitých druhů - šelmy (skot) kopytníci. U hmyzu existuje schopnost svalových
vláken reagovat na nervové impulsy i postupným stahováním.
Inervace - odstředivě - nervosvalová ploténka.
Jedno svalové vlákno je inervováno jediný neuronem,
ale jeden neuron může inervovat více svalových vláken. Odpověď vlákna „vše
nebo nic“. Odpověď svalu (více
neuronů) - postupné stahování a uvolňování. Zpráva o situaci ve svalstvu -
dostředivě - nervosvalová zakončení, šlachová vřeténka (proti přetížení).
Vznik žíhané svaloviny – z embryonálního mezenchymu vznikají tzv. myoblasty, které se spojují v myotuby a
ty postupně diferencují ve svalové vlákno.
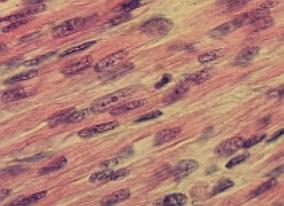
Myotuby
kosterní svaloviny plodu člověka se spojují ve svalová vlákna.
6.3.1. Elektrická tkáň (URL 26)
Jde o zvláštní modifikaci žíhané
svaloviny. Nachází se u tzv. elektrických orgánů některých ryb - elektrický
úhoř
(Electrophorus – URL 24 ), el.
rejnoka (Torpedo), el. sumec (Malopterurus).
Tkáň (URL
25) je tvořena destičkovitými útvary - elektroplaxy, které jsou uspořádány
do sloupce, čímž se sčítá jejich napětí. Jedna strana je bohatě inervována -
akční potenciál 1 ploténky = 0,14.
Úhoř - podél páteře 5 - 6 tisíc
elektroplaxů (plotének) = výboj 600V. Pseudoelektrické orgány - u
hlubinných ryb - přeměna
ocasní okohybné nebo žaberní svaloviny
= série krátkých jemných
elektr. impulsů - komunikace,
orientace.
![]()
6.4. Myokard - srdeční svalovina obratlovců,
liší se od žíhané svaloviny:
a) stavební jednotkou je jednojaderná
buňka - cylindrický útvar, rozvětvená do kontaktu s ostatními, tvoří spolu
složitou protoplazmou síť - plasmodium syncytiální
b) jádra ve středu svalových buněk, ne
pod sarkolemou,
c) sarkoplazma mnohem bohatší na sarkozómy
a chudší na myofibrily. Myofibrily splývají v pentlicovité myotenie (na příčném
řezu v tradiční histologii),
d)
interkalární disky - příčné „přepážky“ mezi buňkami a vlákny probíhají
schodovitě nikoli napříč kolmo. Jde o buněčné kontakty srdeční svaloviny –
desmozómy spolu s nexusovými spoji , snížený elektr. odpor, předávání
podráždění.
e) neexistují regenerační schopnosti
(myokard neschopen regenerace)
Endomysium - retikulární a jemné
kolagenní vazivo, s tukovými buňkami, řečiště kapilár a lymfatických cév.
Autonomní
převodní systém regulující činnost - Keith-Flackův, Aschoft-Tawarův, Hisův
svazek (Purkyňova vlákna).
Nelze
ovlivnit centrálním nervstvem, pouze částečně ovlivnitelné autonomním
nervstvem: parasympatikus – zpomaluje, sympatikus – zrychluje
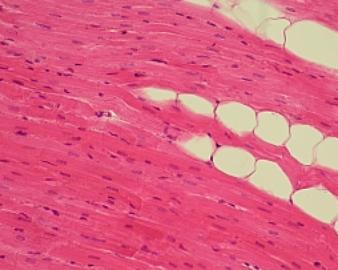
Myokard
při menším zvětšení, svalové buňky prostoupené jemnými vrstvami endomysia ,
které místy vytváří tukové pojivo. VP

Myokard
při větším zvětšení – svalové buňky s protáhlými jádry. Buňky jsou navzájem
propojeny nepravidelnými jemnými přepážkami, tzv. interkalárními disky (např.
v pravém rohu dole). Vlevo dole je světlý prostor adipocytu (tuková buňka
a přibližně uprostřed snímku napříč jemný pruh ŘVP – endomysium
s fibrobcyty a lymfocyty.
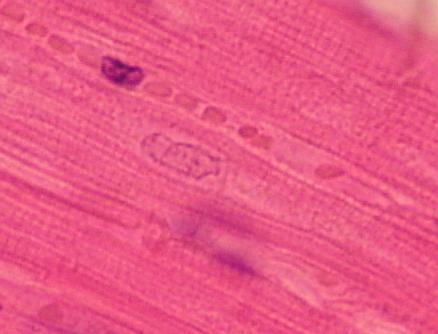
Detail buňky myokardu (světlejší jádro uprostřed), nahoře
zřetelná hustá síť kapilár, které myokard vyživují. Každá z kapilár je
právě tak na objem jediného erytrocytu, které se uvnitř kapilár nacházejí.Tmavé
menší jádro patří některé z buněk endomysia prostupujícího myokard
(vaskularizace, ochrana)
![]()
7. TKÁNĚ NERVOVÉ
- specializované pro vzrušivost a dráždivost
- přenos podráždění
1) s cílem analýzy podnětu a
odpovědi na něj
2) též koordinace činnosti všech
tkání a orgánů hloubkové smysly)
3) registrace procházejících impulzů
- paměti - uloženy
záznamy za celé uplynulé období života
protože nervové buňky
se nedělí, jsou celý
život tytéž - orientace živočichů nejen
v prostoru, ale i v čase. (Teorie
morfické rezonance – nervová soustava pouze jako orgán registrující informace
uložené v poli mimo tělo – R. Sheldrake.)
4) nervová tkáň - tkáň řídící,
spojující všechny buňky v těle v jediný harmonický celek (spolu s
hormony).
Mozek (a nebo) mozkové uzliny - sídlo psychických
projevů.
Specifická
stavba nervové tkáně - vláknitý charakter (průnik do všech částí těla): buňky
nervové, buňky gliové (vyživovací, podpůrné), dále se účastní vláknité
pojivo, cévy.
7.1. Základní systémy nervové tkáně
Systém centrální a periferní
- mozek výrazně vláknitý charakter
-mícha jde
o výběžky nervových buněk
-nervové uzliny (ganglia) nebo jejich svazky = nervy
( těla nervových buněk, = neobsahují nervové buňky
jejich výběžky, mají gliové buňky a jsou
podpůrné a vyživovací složky) prokrvovány
Periferní nervové systémy spojují smyslové
orgány (receptory)
s nervovými centry a centra s tkáněmi
(efektory) (svalové a žlaznaté)
7.2. Nervová buňka - neuron

Schéma neuronu (Janquiera a kol.)

Tělo neuronu – rekonstrukce podle
snímků EM
Gangliová buňka
(cyton, perikaryon) - kulovitá, slzovitá, vřetenitá, polygonální, 10-500 um.
Jádro - kulovité, 1 i více
jadérek
Cytoplazma (neuroplazma)
poměrně málo mitochondrií (podélné přepážky).
Endoplazmatické retikulum - drsné, velmi vyvinuté, na mnoha místech
zduřeno v uzlíky = Nisslova tělíska (tigroidní substance) - nejde do axonu,ano
do
do dendritů (hladké). Nisslova tělíska
= ztluštělé membrány endoplazmatického retikula se zrny 10-30 nm - kvanta RNK a
bílkovin. Vymizí-li z buňky, její nervová aktivita klesá.
Tuky - dosti v
cytoplazmě (lipozómy)
Fe - obsah dosti vysoký (z anorg.
látek)
Na povrchu cytonu - dvojitá membrána =
neurilema.
Neurofibrily - v
cytoplazmě nervových buněk. Podílejí se na šíření nervových vzruchů a vedení
sekretů. Pokračují i ve výběžcích nervových buněk (mikrotubuly a intermediální
filamenta).
Výběžky nervových buněk - dendrity a
neurity
Dendrity - krátké a složitě
větvené (nebo dlouhé), vedoucí vzruchy ze smyslových orgánů = dendrity
axonálního typu. Povrch kryt neurilemou (jen, bez pochev), v bázi Nisslova
tělíska. Vzruchy vedou (dostředivě) směrem do buňky (cellulipetálně).
Neurity
(axony uvnitř, někdy jen axony – nahé neurity bez pochev) - mnohem delší než
dendrity, u člověka až 1 metr. Šířka 1- 4um. U kroužkovců a hlavonožců obrovské
axony o průměru několik um. Bazální část není rozšířena a neobsahuje Nisslova
tělíska. Vedou vzruchy vždy od buňky (cellulifugálně).
Každý
cyton má jdiný 1 axon.
Axon může vytvářet postranní větve
- kolaterály. Na konci jsou neurity i kolaterály rozvětvené v trsy vlákének telodendrony
(telodendrie = jednotlivá ploténka telodendronu), synaptické uzlíky =
konce telodendrií.
Axony - pokryv:
1) nahé - jen axolema
2) pochvy (u obratlovců):
a) Schwannova – tělo Schwannovy buňky tvoří buď myelinovou
pochvou nebo jen kryje část axonu.
b) myelinová (polotekutá, bělavá tukovitá látka,
lecitiny, fosfolipidy, cholesterol, cerebrosidy + skleroproteiny - mnohonásobně obtočené cytoplazmatické
membrány Schwannových buněk.
Schmidt-Landermannovy náručky v mylinové pochvě (zbytky cytoplazmy
Schw. buněk) - neurokeratinová vlákna - matrix myelinové pochvy.
Ranvierův zářez - mezi dvěma sousedními Schwan.
buňkami, membrána Schwan. buněk se dotýká axolemy.
Interanulární nodulus - úsek mezi
sousedními Ranv. zářezy vymezený 1 Schwan. buňkou = internodium.
Axony u bezobratlých, dále všechny
dendrity výstupové a vstupové konce všech axonů a axony čichových nervů
obratlovců jsou nahé, kryté axolemou.
Myelinová a Schwannova pochva se
vyskytuje u většina axonů obvodních nervů obratlovců kromě kruhoústých a
čichových nervů.
Vegetativní nervy všech obratlovců +
axony některých bezobratlých, ryb, kruhoústých mají nezřetelně vyvinutou
myelinovou pochvu a dobře pochvu Schwannovu.
Axony bílé
hmoty mozkové a míšní mají jen myelinovou hmotu.
Souborně viz tabulka:
A X O N Y
|
Nahé |
Myelinová + Schwannova
pochva |
Slabá myelinová + dobrá Schwan. pochva |
Jen
myelinová pochva |
|
-
Bezobratlí (téměř všichni) -
čichové nervy obratlovců -
vstupní a výstupní část axonů
a kolaterál obecně |
- většina
axonů bvodních nervů
Vertebrat kromě Cyclostomat |
-
vegetativní nervy obratl. +
axony kruhoústých , ryb a
některých bezobratlých |
-
axony bílé hmoty mozkové
a míšní |
Všechny dendrity jsou nahé
Neurony (podle tvaru
gangliových buněk) lze rozdělit na:
1)
Unipolární (adendritické) - oválný tvar a jen jeden neurit
(ganglion hlemýždě),
2)
zdánlivě unipolární - 1 dentrit a 1 neurit,
3)
bipolární - vřetenovitý tvar, na jednom pólu dendrit a na
jiném neurit,
4)
multipolární - hvězdicovité, 1 neurit a vícero dendritů.
Podle délky axonů:
- dlouhoaxonové neurony (Deitersovy
buňky) - dlouhé neurity od periferního nervstva,
- krátkoaxonové (interkalární) neurony
- např. v kůře mozečku, Purkyňovy buňky.
7.3. Spojení
nervových buněk
Prostřednictvím
nervového zápoje = synapse (presynaptický a postsynaptický
neuron) - podle směru šíření vzruchu. Zápoje: axodendritické,
axoaxonické - na bázi axonu postsynaptického neuronu, axosomatické - na povrch
buňky nervové, svalové, žlázové. Synapse tvoří:
1.
Synaptický uzlík
2.
Intersynaptický prostor (asi 12 nm)
3.
Mediátory - vlastní přenos nervového vzruchu. (Mediátorové váčky v
uzlíkovitých rozšířeninách telodendrií.
Nervová zakončení: cholinergní
(acetylcholin)
adrenergní
(noradrenalin)
U bezobratlých serotonin, dopanin,
gama-aminomáselná kys.
V okamžiku proniknutí vzruchu do
synaptického uzlíku se mediátor vyleje do intersynaptického prostoru. Kontakt
mediátoru s membránou postsynaptické buňky vyvolá vzruch v cílové buňce.
Vedení vzruchu - membránový
potenciál: klidový, akční.
Saltatonní vedení vzruchu pouze na
místě Ranvierových zářezů. (Na+ se mění s K+
a
pak zase zpět za vynaložení energie).
Rekonstrukce synapse podle EM
Biochemismus
synaptických reakcí – mediátory musejí být vzápětí rozloženy, aby vzruch ustal.
Prekurzory mediátorů se znova
v synaptickém knoflíku regenerují. (Janquiera a kol.)
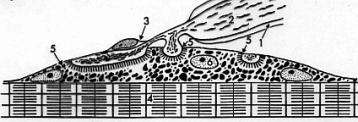
Schéma
nervosvalové ploténky (rekonstrukce podle EM, (Klika a kol.): 3 – nervové
zakončení, 5 synapse, 6 jádra svalového vlákna
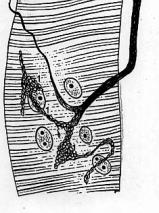
Nervosvalová
ploténka, pohled zevně svalového vlákna. (Klika a kol.)
Gliové buňky mají různé
funkce i tvar.
1) Makroglia - velké gliové
buňky:
a) ependymové
- cylindrické na bázi s kořenovitým výběžkem. - Výstelky mozkových dutin a
míšního kanálu. Kinocilie pohybují míšním mokem v dutinách, kořeny proplétají
nervovou tkáň. Evertované žlázy (z ependymu) - plexus chorioideus - bílkovinná
složka mozkomíšního moku.
b) astrocyty - hvězdicovitě
větvené buňky s kořenovitými výběžky, perivasculátní nožka.
2)
Oligodendroglia - podobné astrocytům, jsou však menší a mají méně
výběžků. V bílé hmotě mozku a míchy a v a synaptických uzlinách podél míchy.
Oporná funkce, vytváří myelinové pochvy (patří se i buňka Schwannovy).
Vlevo dva
astrocyty s perivaskulární nožkou, vpravo nahoře olgodendroglie a dole
mikroglie (makrofág)
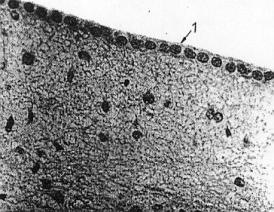
Ependym
(výstelka mozkových komor). Buňky mají ještě výběžky do mozkové tkáně (zde
nejsou zřetelné. (Klika a kol., 1986)
3) Mikroglia - nejmenší
buňky se schopností amoeboidního pohybu
= jediné volné buňky v nervové tkáni. Ochrana, fagocytóza (makrofágové –
v paradoxu ke svému názvu).
Mikroglie
(Klika a kol., 1986,)
7.4.
Diferenciace nervové tkáně
Z
ektoblastu (ale již asi změna názoru) se vyvíjejí tzv. neuroglioblasty a
z nich pak neuroblasty a spongioblasty. Z neuroblastů = neurony,
ze spongioblastů = buňky gliové. Neuroblasty, spongioblasty i gliové
buňky - mají schopnost dělení. Neurony se již nedělí. Velmi slabá regenerace
nervových buněk. Axony prorůstají do pochev, jsou li tyto zachovány.
Nervy
i centrální nervový systém (mozek, mícha, uzliny, nervy) jsou tvořeny nervovými
buňkami, gliovými buňkami a pojivem různého typu, které umožňuje vaskularizaci,
ochranu a zpevnění. Mezi krevním systémem a nervovými buňkami existuje bariéra.
Tu překonávají neuroglie tím, že z cév přebírají živiny a předávají je
neuronům.
Příklad stavby orgánu – nervu:
Nad
Schwannovou pochvou se nachází jemné endoneurium z vláknitého pojiva
(bazální membrána). Několik neuritů pak obaluje perineurium -
pojivový obal. Všechny obaly jsou pospojovány navzájem - plsťovitá výztuž
(mechanická ochrana nervů) a navenek obklopeny vazivovým pouzdrem, jemuž se
říká epineurium a které obaluje vzniklý nerv. Nervy jsou tedy komplexy
nervové tkáně a pojiv.

Nerv
z různými typy neuritů (Klika a kol., 1986) Nahoře a dole dva neurity
s myelinovou (tmavé mezikruží) i Schwannovou pochvou (na povrchu), 3 –
jádro Schwannovy buňky. Uprostřed jsou neurony s pochvou myelinovou. 4 –
jádro neurogliové buňky. Axony jsou jakoby fagocytovány buňkami, které je
obalují. Světle šedé amorfní plochy mezi neurity jsou okrsky endoneuria (Klika
a kol, 1986).
Spinální nervová uzlina kočky – velké buňky s růžově
zbarvenou cytoplasmou a velkými, výraznými jádry – neurony - jsou obklopeny
neurogliemi (malá jádra kolem). Vše je drženo pohromadě vláknitým pojivem –
uvnitř řídké, na povrchu uzliny husté.
![]()
8. POHLAVNÍ BUŇKY
8.1. Typy gamet
Gamety : samčí -
pohyblivé
samičí - nepohyblivé
Izogamety - stejné
anizogamety - různé
mikrogamety - samčí
makrogamety – samičí
8.2. Vznik gamet - z tzv.
primordiálních gonocytů (prapohloavních buněk), které v rané fázi
embryonálního vývoje si organismy uschovají na různých místech zárodku (pro
každý druh typické) a které později migrují do vyvíjejících se pohlavních
orgánů, kde se usazují. Pojivo pohlavních orgánů a buňky zárodečných epitelů
přebírají o prapohlavní buňky péči. Prapohlavní buňky podle hormonální regulace
procházejí následujícími fázemi:
1) rozmnožovací - série mitotických dělení, která dá velký
počet tzv. gametogonií (spermatogonie nebo oogonie). V danou dobu se
gametogonie přestanou mitoticky dělit a vstupují do fáze
2) růstové – Zvětšuje se objem
cytoplazmy (zvláště u samičích buněk – žloutek). Buňky se mění v tzv.
gametocyty (spermatocyty nebo oocyty) prvního řádu (gametocyty I). Gametocyty I
vstoupí do prvního zracího dělení. Proděláním meiózy I vzniknou gametocyty II, ty
druhým meiotickým dělením dají vznik ootidě a 2-3 pólovým buňkám nebo 4
spermatidám (viz meiosis v cytologii).
3) zrací – spermatidy
prodělávají spermateliózu, - tvoří se bičík, ztrácejí, cytoplazmu, vytvoří se
perforatorium a akrozóm v přední části, jádro klidové. (Ootida prodělá
tuto fázi často až po oplození ve vejcovodech matky - jde v podstatě o
dokončení 2. meiotického dělení a vznik haploidního jádra.)
9. Použité www stránky:
1. http://www.ucmp.berkeley.edu/porifera/porifera.html
2. http://www.bu.edu/histology/p/20402loa.htm
3. http://www.ulb.ac.be/sciences/biodic/ImMembrane.html
4. http://www.bioscience.org/atlases/tumpath/em/desmosom.htm
5. http://www.apsnet.org/education/IllustratedGlossary/PhotosN-R/plasmodesma.htm
6. http://www.mun.ca/biology/desmid/brian/BIOL2060_W2003/CellBiol11/CB11_25.html
7. http://www.unm.edu/~leonardm/devel.html
8. http://www2.uerj.br/~micron/atlas/atlasenglish/tissue.htm
9. http://www.botany.uwc.ac.za/SCI_ED/grade10/mammal/Epithelial.htm
10. http://www.scientia.org/cadonline/Biology/specialcells/epithelial.ASP
11. http://www.unomaha.edu/~swick/2740epithelium.html
12. http://web.bio.utk.edu/kennedy/GlandularEp_files/frame.htm
13. http://www.usc.edu/hsc/dental/ghisto/epi/
14. http://www.ulb.ac.be/sciences/biodic/ImAnatepon0002.html
15. http://www.bethel.edu/~kisrob/bio113/ClassSessions/10_5_01/10_05_01.html
16. http://www.science.ubc.ca/~biomania/tutorial/tuthisto/ep04.htm
17. http://www.bioeng.auckland.ac.nz/anatml/anatml/database/cells/cells/parts/part/part_33.html
18. http://cellbio.utmb.edu/microanatomy/epithelia/epith_lec.htm#Stratified%20epithelia
19. http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/C/Collagens.html
20. http://www.esg.montana.edu/esg/kla/ta/tissues.html
21. http://meds.queensu.ca/medicine/anatcell/phaseone/page14.htm
22. http://www.meddean.luc.edu/lumen/MedEd/Histo/frames/histo_frames.html
24. http://www.flmnh.ufl.edu/fish/tropical/JSA/gymno.htm
25. http://helium.vancouver.wsu.edu/~ingalls/eels/Electric_Anatomy/electric_anatomy.html
26. http://www.sbg.ac.at/ipk/avstudio/pierofun/ray/eod.htm
27. http://www.okc.cc.ok.us/biologylabs/Documents/Porifera_Cnidaria/Hydra_longitudinal.htm
28. http://www.vcbio.sci.kun.nl/eng/image-gallery/show/labels/print/AN0014/
29. http://www.histol.chuvashia.com/atlas-en/digestive-05-en.htm
30.
http://www.rkm.com.au/CELL/obesity-adipocytes.html
31.
http://www.diaglab.vet.cornell.edu/clinpath/modules/heme1/intro.htm
10. Použitá literatura
Dogiel V. A., 1961: Zoologie Bezobratlých, SPN Praha, 598 s.
Jauquiera L.C., Carneiro J., Kelly R. O.: Základy histologie,
Nakladatelství a vydavatelství H & H, 1999, 502 s
Klika E., Vacek Z, Dvořák M., Kapler K. (1986): Histologie. Avicenum
zdravotnické nakladatelství Praha. 612 s.
Knoz
J.: Obecná zoologie, I. Taxonomie,
látkové složení, cytologie a histologie.
II. Organologie, rozmnožování,
vývoj
živočichů a základy biologie. Skripta
PF UJEP v Brně, 3. vydání, 1984, 340 s.